HÜCRE BİYOLOJİSİ
DERS NOTLARI
Doç. Dr. Nurettin YÖREK
1
Hücrenin Temel Yapı Taşları ve Kimyasal Bileşimi
Bir hücre içindeki bileşiklerin % 99’undan fazlası başlıca altı elementten meydana gelmiştir.
Bu elementler karbon (C), hidrojen (H), nitrojen (N), oksijen (O), fosfor (P) ve kükürt (S)’dür.
Karbon ile diğer elementler arasında kurulan kovalent bağlar sonucunda çok sayıda bileşik
meydana gelebilir. Nitekim hücre içerisinde molekül ağırlıkları 100-1000 arasında değişen
yaklaşık 1000 çeşit molekül bulunur. Bu moleküller stoplazmada çözünmüş olarak yer alırlar.
Çeşit olarak çok sayıda olmakla beraber hücrenin kimyasal yapısında yer alan bu organik
molekülleri dört ana grupta toplamak mümkündür. Bunlar: karbonhidratlar, yağlar,
aminoasitler (proteinler) ve nükleotidlerdir. Bu dört gurup organik bileşiğe hücre ağırlığının
% 70-80 kadarını kapsayan su ve dış ortamdan alınan elektrolitler de eklenince hücrenin
kimyasal yapısı ortaya çıkar.
Su: Hücrenin temel maddesi sudur diyebiliriz. Nitekim hücre ağırlığının % 70-80 kadarını su
oluşturur. Hücre içi reaksiyorların büyük kısmı su ortamı içerisinde yer alır. Su ayrıca
maddelerin difüzyonlarına veya bir yerden diğerine akışına olanak sağladığı için hücre içi
taşımanın gerçekleştirilmesini sağlar. Su aynı zamanda diğer kimyasal maddelerin davranışını
belirleyen çok önemli bazı fiziksel özellikler taşır. Su molekülü aynı sayıda elektron ve proton
taşıdığı için nötr bir moleküldür. Su yüksüz olmakla birlikte taşıdığı elektronların dağılımı
dengesizdir ve bu nedenle molekülde bir kutuplaşma ortaya çıkar. Oksijen atomunun
çekirdeği 8 proton taşır ve hidrojen atomlarının elektronlarını kendine doğru çeker. Bu yüzden
su molekülünde oksijen atomunun bulunduğu bölge, hidrojen atomlarının bulunduğu bölgeye
oranla biraz daha negatiflik kazanır. Buna karşın hidrojen atomlarının bulunduğu bölge
hafifçe elektropozitiftir. Su katı madde molekülleri ile zayıf elektiriksel bağlar kurar. Bu
özelliği nedeniyle su birçok katı madde için çözücü görevi yapar. Bunun tipik bir örneği
NaCl’ün su içerisinde çözünmesidir. Zıt yüklü olan Na ve Cl atomları arasındaki kuvvetli
çekim nedeniyle katı NaCl kristalleri oluşur. Buna karşın su
molekülleri Na+ ve Cl- iyonlarına doğru çekilirler ve bu iyonların çevresi su molekülleri ile
kaplanır. Sonuçta iyonların etrafında su moleküllerinden oluşan bir tabaka oluşur. Buna
iyonların “hidrasyon kabuğu” adı verilir. Etraflarında hidrasyon kabuğu olan Na+ ve Cliyonları arasındaki çekim gücü zayıflar ve iyonlar birbirlerinden ayrılırlar. Böylece NaCl su
içinde eriyik halinde bulunur. İyonların çevresindeki bu hidrasyon kabuğu bazı iyonların
hücre membranındaki porlardan geçişlerini etkileyen önemli bir faktördür. Yapısında iyonik
ya da polar guruplar taşımayan moleküller ise suda çözünmezler. Su çözücü fonksiyonunun
yanı sıra bazı reaksiyonlara girebilir veya bir reaksiyon sonucu açığa çıkabilir.
Karbonhidratlar: Karbonhidratlar hücrede yapısal değil fonksiyonel olarak işlev görürler.
Bazı karbonhidratlar hücredeki (özellikle plazma membranındaki) lipitlerin ve proteinlerin
yapısına girerek glikolipit ve glikoproteinleri oluştururlar. Glikolipit ve glikoprotein
molekülleri yapısal olarak çok genişbir çeşitlilik gösterirler ve bu möleküller hücrelerin
birbirlerini tanıma olayında önemli roller oynarlar. Fonksiyonel olarak karbonhidratların
hücre için önemi besin maddesi olarak enerji eldesinde kullanılmalarıdır. Bu açıdan glukoz
pek çok hücrenin temel besin kaynağını oluşturur ve glukozun bir dizi oksidatif reaksiyonlar
sonucu CO2 ve suya kadar yıkımı sonucunda hücrenin fonksiyonlarını yerine getirebilmesi
için gerekli olan enerji (ATP) elde edilir. Bir kısım hücreler gerektiğinde kullanmak üzere
glukozu bir glukoz polimeri olan glikojen halinde depo edebilirler.
Yağlar: Yağların hücrede hem yapısal hem de fonksiyonel görevleri vardır. Yağlar toplam
hücre kitlesinin % 15 kadarını oluştururlar. Fosfolipitler ve kollestrol gerek plazma
membranının gerekse hücreiçi organalleri kuşatan membranların yapı taşlarıdır. Buna ek
olarak trigliseritler (nötral yağlar) hücre için önemli bir yakıt kaynağıdır ve bunların
2
yıkımından enerji elde edilir. Trigliseritler bazı özel hücrelerde, örneğin yağ hücrelerinde,
hücre kütlesinin % 95’ini oluşturacak derecede depo edilebilirler.
Proteinler: Hücre kütlesinin %10-20’sini proteinler oluşturur. Hücrede yapısal ve globüler
olmak olmak üzere iki tip protein bulunur. Genellikle ince flamentler halinde yeralan yapısal
proteinlerin fonksiyonları arasında en önemlisi kas kasılmasını sağlamaktır. Buna ek olarak
filament halindeki bu tip proteinler siller ve mitotik iğ iplikçikleri gibi organellerin yapısını
oluşturan mikrotubuller halinde de organize olmuşlardır. Bu örneklere hücrelerarası
haberleşme bölgelerinin (gap junctions) yapısını oluşturan proteinler de eklenebilir. Globuler
proteinler ise filament değil, küresel yapıdadırlar ve bunlar esas olarak hücrenin enzimleridir.
Bu tip proteinler hücre sıvısı içinde çözünmüş veya membranların yüzeylerine tutunmuş veya
membranlara inkorpore olmuş durumda bulunurlar. Enzimler hücredeki diğer maddelerle
doğrudan temas ederek kimyasal reaksiyonları kataliz ederler. Böylece enzim proteinleri
hücrenin metabolik fonksiyonlarını da düzenler.
Nükleotidler: Bir nükleotid bir baz, bir şeker ve bir fosfat molekülünden oluşur. Bazlar
nitrojen taşıyan, halka yapısında moleküllerdir ve beş karbonlu bir şekere (riboz veya
deoksiriboz) bağlanırlar. Bu şekere en az bir tane de fosfat grubununu bağlanması ile bir
nükleotid oluşur. Bazlar pirimidinler ve pürinler olmak üzere iki grubu ayrılırlar. Pirimidinler
sitozin, timin ve urasil, pürinler ise adenin ve guanindir. Nükleotidler hem kimyasal enerji
taşıyıcıları, hem de genetik bilgi depoları olan nükleik asitlerin yapı taşlarıdır. Adeninin bir
trifosfat esteri olan adenozintrifosfat (ATP) evrensel bir eneji kaynağı olarak hemen bütün
biyokimyasal reaksiyonlar için gerekli enerjiyi sağlar. Özel bir nükleotit,
siklikadenozinmonofosfat (cAMP) bir hücre içi haberci sistemi oluşturarak pekçok hücre içi
aktiviteyi kontrol eder. Nükleotidlerin çok özel bir rolü de nükleik asitlerin yapıtaşlarını
oluşturmalarıdır. Burada bir nükleotitin şeker molekülündeki 3’-hidroksil grubu diğer bir
nükleotitin 5’-fosfat grubu ile bağ yaparak nükleik asit zincirlerini meydana getirirler Başlıca
3
iki tip nükleik asit vardır Bunlardan ribonükleik asit (RNA) de şeker grubu bir ribozdur.
Deoksiribonükleik asit (DNA) de ise ribozun 2’ pozisyonundaki hidroksil grubu yerini bir
hidrojene bırakmıştır (deoksiriboz). RNA’da yer alan bazlar adenin, urasil, guanin, ve
sitozindir. DNA da ise urasilin yerine timin bazı da yer alır. Bu iki nükleik asitin baz
dizilişleri hücrenin genetik bilgisini temsil eder.
Elektrolitler: En önemli hücre içi elektrolitleri potasyum (K+), magnezyum (Mg+2), fosfat,
sülfat ve bikarbonattır. Bunlara ek olarak, az miktarlarda olmak üzere, sodyum (Na+), klor
(Cl-) ve kalsiyim (Ca+2) de sayılabilir. Elektrolitler hücre suyunda çözünürler ve hücresel
reaksiyonlara katılan inorganik kimyasal maddeleri de meydana getiriler. Bazı hücresel
kontrol mekanızmalarının işlemesinde de elektrolitler rol oynarlar. Örneğin, Na+ ve K+ sinir
ve kas hücresi gibi uyarılabilen hücrelerde elektrokimyasal impulsların iletimini sağlar.
Birçok hücre içi elektroliti kofaktör görevi görerek enzimatik kataliz reaksiyonlarında rol
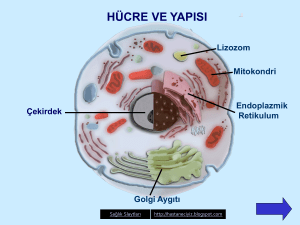
HAYVAN HÜCRESİ: GENEL YAPI VE ORGANELLERİ
Hayvan vücudunda yer alan organlar hücrelerarası destek sistemleri aracılığı ile birarada
tutulan çok sayıda hücredenoluşmuşlardır. Farklı organların yapısını meydana getiren hücreler
farklı görevler yapmak üzere özelleşmeşlerdir. Bu hücreler birbirlerinden yapısal ve
fonksiyonel olarak farklı iseler de hepsinin ortak bazı temel karakteristikleri vardır. Örneğin
her hücre yaşamını sürdürebilmek için beslenmek zorundadır ve bütün hücreler hemen
tamamen aynı tip besinleri kullanırlar. Bütün hücreler enerji eldesi için oksijene
bağımladırlar. Besin maddelerini enerjiye çevirme mekanizması tüm hücrelerde temelde
aynıdır ve tüm hücreler metabolizmaları sonucunda oluşan artık maddeleri kendilerini
çevreleyen sıvılara bırakırlar. Görüldüğü gibi hücre sadece hücre sıvısı, enzimler ve diğer
kimyasal maddelerden oluşan bir yapı olmayıp ileri düzeyde organize olmuş, organel adı
verilen fiziksel yapılar da taşımaktadır. Bu organellerin herbiri farklı bir fonksiyonu yerine
getirmek üzere özgülleşmişleridir. Örneğin, lizozom hücre içi sindirim, mitokondri solunum,
endoplazmik retikulum ve golgi aparatı sekresyon, çekirdek nükleik asit sentezi
fonksiyonlarından sorulmudur. Bir membranla çevrilmiş olan bu organellerin varlığı bir
okaryotu prokaryottan ayıran en önemli farklılıktır. Bu derste bir ökaryotik hücredeki
organellerin yapı ve fonksiyonlarını inceleyeceğiz.
Hücre (plazma) membranı
Bütün hücreler, genellikle plazma membranı olarak anılan bir sitoplazmik membran ile
karakterizedir. Esas olarak protein ve lipidlerden oluşan bu membranın, hücrenin yapısal
bütünlüğünü koruyan fiziksel bir bariyer olma görevine ek olarak, hücrenin yaşamını
sürdürebilmesi için gerekli olan bir takım fonksiyonları da vardır. Bu fonksiyonlar arasında,
zararlı atık maddelerin hücre dışına atılması, besin maddeleri ve enerji kaynaklarının hücre
içine alınması, çoğalma haraket etme ve çevredeki bileşenlerle etkileşim kurma gibi olaylar
sayılabilir. Biyolojik
membranların taşıdıkları pompalar ve geçitlerden ileri geler yüksek düzeyde bir seçici
geçirgenlik özellikleri vardır. Bu tarnsport sistemleri hücre içi ortamın moleküler ve iyonik
bileşimini üzenler.
Membranlar hücreler ile çevreleri arasındaki bilgi akışını da kontrol ederler. Dışarıdan gelen
uyarımları alabilmek için özgül reseptörler taşırlar. Bakterilerin besinlerine doğru haraket
etmeleri, hedef hücrelerin hormonlara karşı verdikleri cevap, ışığın algılanması gibi olaylarda
ilk meydana gelen işlem sinyalin membrandaki özgül bir reseptör tarafından alınmasıdır. Bazı
membranlar ise kimyasal ve elektriksel olabilen sinyaller üretirler. Böylece membranlar
biyolojik bilgi alış-verişinde önemli bir rol oynarlar. Biyolojik sistemlerde iki önemli enerji
4
dönüşüm olayı, yüksek düzeyde organize olmuş enzimleri ve diğer proteinleri taşıyan
membranlarda meydana gelir.
Membranların ortak özellikleri: Membranlar fonksiyonları bakamından olduğu gibi yapları
bakımından da birbirlerinden farkılılık gösterirler. Bununla birlikte bazı önemli ortak
özellikler de taşırlar. Bu ortak özellikler şu şekilde sıralanabilir:
1. Membranlar birkaç molekül kalanlığında ince tabaka yapısındadır. Birçok membranın
kalınlığı, 60-100 A arasındadır.
2. Esas yapılarını protein ve lipidler oluşturur. Membranlarda ayrıca protein ve liptlere bağlı
karbonhidratlar da bulunur.
3. Membran lipitleri, hem hidrofilikhem de hedrofobik kısımları bulunan nispetin küçük
moleküllerdir. Bu lipitler sulu bir ortamda kendiliklerinden kapalı iki tabakalı yapılar
oluştururlar. Bu lipit tabakaları yüklü moleküllerin geçişi için bariyer görevi görürler.
4. Özgül proteinler membranın belirli fonksiyonlarına aracalık ederler. Membran proteinleri,
pompa, geçit, reseptör, enerji transdüseri ve enzim olarak fonksiyon görürler. Bu proteinler
lipit tabakaları arasında yer alırlar ki, bu liptler proteinlerin görev yapabilmeleri için uygun
ortamı sağlarlar.
5. Membranlar nonkovalent yapılardır. Membranların yapısını oluşturan protein ve lipit
molekülleri birçok nonkovalent bağlantılar kurmuşlardır.
6. Membranlar asimetriktir. Membranların iç ve dış yüzeyleri birbirlerinden farklıdır.
7. Membranlar sıvı yapıdadır. Lipit yapılar membranın yüzeyine difüze olmuşlardır. Özgül
birtakım bağlantılar yapmadıkları zaman proteinler de bu şekilde yerleşmişlerdir. Membranlar
“proteinlerin ve liptlerin iki, boyutlu çözeltileri olarak tanımlanabilir.
Sıvı Mozaik Modeli
Biyolojik membranların yapıları ile ilgili olarak buraya kadar yazdıklarımızı toparlayacak
olursak ortaya yeni bir membran modeli çıkar. Bu modeli ilk defa 1972 yılında Singer ve
Nicolson ortaya koyarak adına “sıvı mozaik modeli” dediler ve membranları globuler
proteinlerin ve lipitlerin iki-boyutlu çözeltileri olarak tanımladılar ( şekil 4. 22.). Sıvı mozaik
modelin en belli başlı özelliklerini şöyle özetleyebiliriz:
1. Membrandaki fosfolipitler ve glikolipitler bir çift-tabaka ( bi-layer) oluştururlar. Bu çift
tabaka hem bir geçirgenlik seddi, hemde integral proteinler için çözücü ortam fonksiyonu
görür.
2. Membran lipitlerinin küçük bir kısmı membrandaki proteinler ile özgül bağlantılar
kurmuşlardır ve bu bağlantılar proteinlerin fonksiyonları için gereklidir.
3. Membran lipitleri ve proteinleri membran içinde lateral difüzyon yapabilirler: fakat
proteinler membranın bir yüzeyinden diğerine geçemezler.
4. Membranlar asimetriktir. Bu asimetriyi sağlayan başlıca faktörler membran proteinleri ve
karbohidratlarıdır.
5
Küçük Moleküllerin Membranda Transportu
Küçük moleküllerin membranları geçişleri başlıca iki yolla olur. Bunlar:
(1) Difüzyon, (2) Aktif transporttur.
Difüzyon
Molekül, iyon gibi taneciklerin yüksek konsantrasyon bölgelerinden düşük konsantrasyon
bölgelerine doğru net akışına difüzyon denir. Bir molekül ne kadar küçük ve yağda
eriyebilirse lipit tabakasını o kadar kolay geçer. Bu tip moleküllere örnek O2 ve N2'dir.
Bunlar hem küçük hem de hidrofobik olduklarından lipit tabakası içinde hızla difüzyon
yaparlar. Benzen molekülü biraz
daha büyük olmasına rağmen hidrofobik özelliğinden dolayı lipit tabakasını kolaylıkla geçer.
Lipit tabakasını net bir elektrik yükü olmayan polar moleküller de geçebilirler. Bu
moleküllerin difüzyonunu sınırlayan faktör bunların büyüklükleridir. Örneğin polar; fakat
küçük moleküllerden CO2, üre ve gliserol lipit tabakasını kolaylıkla geçerler. Lipit tabaksını
en hızlı geçebilen polar molekül sudur. Suyun difüzyon hızı o kadar yüksektir ki, diğer
moleküllerin veya iyonların membrandaki difüzyonları suya göre ifade edilir. Diğer bazı polar
moleküller, örneğin glukoz, sükroz büyüklükleri nedeniyle lipit tabakasından difüzyonla
geçemez. Lipitte difüzyon yönünü membranın iki yüzeyi arasındaki konsantrasyon farkı
belirler ve moleküller yüksek konsantrasyonda düşük konsantrasyona doğru hareket ederler.
Osmos
Farklı iki çözeltiyi ayıran yarı geçirgen bir membrandan su veya başka çözücü moleküllerin
geçmesi difüzyonun özel bir halidir ve osmos olarak adlandırılır. Hücrenin içinde ve dışında
hücre membranının seçici geçirgenlik özelliğinden dolayı bir konsantrasyon farkı vardır. Su
molekülleri derişik tarafa doğru yani su moleküllerinin daha az olduğu tarafa doğru hareket
ederler. Yoğun tarafta artan sıvının oluşturduğu hidrostatik basınç daha fazla su geçişini
engeller ve bir denge kurulur. Yarı geçirgen bir zardan derişik ortama doğru su geçişini
engellemek için uygulanması gereken basınca o ortamın osmotik basıncı adı verilir.
6
Aktif transport
Aktif transportta proteinin aktivitesi için metabolik enerjiye gereksinim vardır. Bunu enerji ya
doğrudan ATP'nin hidrolizinden elde edilir, ya da bir iyon gradiyenti oluşturularak sağlanır.
Bu enerji sayesinde maddeler konsantrasyon gradiyentinin aksi yönünde (düşük
konsantrasyonda yükseğe doğru) taşınabilirler. Aktif transportta çoğu kez bir maddenin
membranda taşınması başka maddenin de aynı anda taşınmasına bağlıdır. Böyle bir sisteme
ko-transport sistemi denir. Bu sistemde her iki madde aynı yöne taşınıyorsa transporta
simport, maddeler zıt yönler taşınıyorsa antiport adı verilir. Bazı membran transport
proteinleri ise maddeleri sadece membranın bir yüzünden diğerine taşırlar ki bu taşımaya da
üniport denir. Üniport örneklerine difüzyonda da rastlanır. Taşıyıcı proteinin ATP
hidrolizinden elde edilen enerjiyi kullandığı bir antiport sistemine en iyi örnek Na+-K+
pompasıdır. Hemen tüm ökaryotik hücrelerin plazma membranında yer alan bu iyon pompası
sinir ve kas gibi uyarılabilen hücrelerdeki membran potansiyelinin oluşmasında hücre
hacminin kontrolünde ve bazı maddelerin hücrelere ko-transportunda önemli roller oynar.
Büyük (makro) Moleküllerin ve Partiküllerin Membranda Transportu
Membran transport proteinlerinin oluşturduğu transport mekanizmaları sadece iyonların ve
küçük polar moleküllerin membrandan geçişlerinin sağlayabilirler. Bu mekanizmalar
makromoleküller denilen proteinlerin, polisakkaritlerin veya polinükleotitlerin membranını
geçmeleri için yetersizdir. Buna karşın hücre membranlarında makromoleküllerin, hatta çeşitli
7
partiküllerin transportu mümkün olmaktadır. Bu transport küçük moleküller ve iyonlar için
olanlardan çok farklı bazı mekanizmalarla gerçekleşir. Bu mekanizmalar sayesinde büyük
moleküller ve partiküller gerek hücre dışından içine, gerekse hücre içinden dışına doğru
hareket edebilirler. Transport hücre içinden dışına doğru yer alıyorsa buna ekzositoz denir.
Transport hücre dışından içine doğru ise endositoz adı verilir.
Hücre içine alınan maddelerin büyüklüklerine göre endositoz ikiye ayrılır. Sıvıların ve sıvı
içinde çözünmüş nispeten küçük maddelerin hücreye alınmalarına "içme" anlamına gelmek
üzere pinositoz, büyük partiküllerin veya hücre kalıntılarının ( bazen bütün bir hücrenin)
hücre içine alınmasına "yeme" anlamında fagositoz adı verilir. Bütün hücreler sürekli olarak
pinositoz yaparlar; fakat fagositoz ancak bu iş için özelleşmiş hücreler tarafından yapılır
(örneğin makrofajlar, parçalı çekirdekli lökositler). Gerek ekzositozun, gerekse endositozun
en belirgin özelliği taşınacak maddelerin membranla çevrili veziküller içine alınmaları ve
böylece diğer hücre içi maddelerle karışmamalarıdır. Fagositoz sırasında oluşan büyük
veziküllere genellikle vakuol adı verilir. Veziküller ekzositozda hücre membranı ile birleşip
kaynaşırlar (füzyon). Bu kaynaşma, henüz bilinmeyen bir mekanizmayla membranın özgül
bölgelerinde meydana gelmektedir.
Ekzositoz
Hücre içinde
sentezlenen çeşitli maddelerin
hücre dışına çıkartılması
olayına sekresyon denir.
Ökaryotik hücrelerin bir çoğu
sekresyon yapabilirler ve
sekresyon hemen her zaman
ekzositoz yolu ile yapılır.
Hücre dışına çıkartılan
maddelerin üç akıbeti vardır.
Bunlar ya ekzositoz oldukları
hücrenin yüzeyine tutunurlar,
ya ekstraselüler matrikste kalırlar ya da kan dolaşımına katılırlar.
Endositoz
Birçok hücrede endositoz sürekli olarak meydana gelir ve bu tip endositozda ekstraselüler
matriks ve bunun içinde çözünmüş maddeler, veziküller içinde hücreye girerler. Buna sıvı-faz
endositozu (pinositoz) denir. Başka bir tip endositoz ise ancak hücre endositoz için uyarıldığı
zaman görülür. Buna reseptör aracılı endositoz denir.
Hücrelerarası Bağlantı Bölgeleri:
Bir dokuyu oluşturan hücrelerin birbirleriyle temas ettikleri bölgelerde, varlıkları ancak
elektron mikroskobu ile saptanabilen bağlantı bölgeleri vardır. Bu bölgelerde karşalıklı gelen
8
plazma membranları, membranların hemen altındaki sitoplazma bölgesi ve hatta hücrelerarası
boşluk özelleşmişlerdir. Hücrelerarası bağlantı bölgeleri başlıca üç ana grupta toplayabiliriz.
Bunlar:
1. Yapıştırıcı bölgeler: Hücreleri mekanik olarak biarada tutan bu bölgelere desmozomlar
denir ve bunlar üç tiptir:
a. Kemer desmozomlar
b. Düğme desmozomlar
c. Hemidesmozomlar
2. Sıkı bağlantı bölgeleri (tight junction): Hücreleri sadece birada tutmakla kalmayıp aynı
zamana küçük moleküllere dahi geçirgenliği olmayan bölgler meydana getirirler.
3. İletişim (geçit) bölgeleri: Yan yana gelen iki hücre arasında küçük moleküllerin ve
iyonların bir hücreden diğerine geçişini sağlayan bölgelerdir. Bunların başlıca iki tipi
şunlardır:
a. Geçit bölgleri (gap junction)
b. Kimyasal sinapsis bölgeleri
Bu bölgelerden, çeşitli fizyoloji kitaplarının sinir fizyolojisi ile ilgilibölümlerinden detaylı
olarak bulabileceğimiz kimyasal sinapsisleri bir tarafa bırakıp diğerlerini inceleyelim.
t.j. tight junction (sıkı bağlantı),
a.b. adhesion belt (kemer desmozom)
d.s. düğme desmozom
g.j. gap junction (geçit bölgeleri)
h.d. hemidesmozome
Desmozomlar:
Desmozomlar hücreleri birada tutarak onların
yapısal bir birim halinde fonkisyon
görmelerini sağlarlar. Çeşitli dokularda
yaygın olarak bulunmakla beraber kalp kası,
deri epitel tabakası ve uterus boynuzu gibi
fiziksel etkilerle karşılaşan dokularda daha
bol olarak bulunurlar. Kemer, düğme ve
hemidesmozom olmak üzere üç tiptirler. Bu
üç tip desmozom birçok epitel hücresinde
birlikte bulunurlar.
Bunlardan kemer desmozomlar yanyana
gelen hücrelerden herbirini çepeçevre saran bantlar halindedirler. İki hücrenin karşılıklı gelen
bantları birbirlerine filament yapısında bazı uzantılar ile bağlıdır. Hücre içnde ise plazma
membranının hemen altında ve bantlar boyunca aktin demetleri uzanır. Aktin flamentelirinin
muhtemel fonksiyonu morfogenezde önemli bir basamak olan epitel tabakasının kıvrılıp tüp
oluşturmasını sağlamaktadır.
9
Düğme desmozomlar yanyana gelen iki hücre
membranında yer alan yapılardır. Görünüş ve
fonksiyon olarak perçin çivlerini andırılar. Hücre
içinde, hücreyi boydan boya kateden ve keratin
filamentleri (veya tonofilamentler) adı verilen
yapılar ile birbirlerine bağlıdırlar. Bu filamentler
sitoplazmada bir ağ yapısı oluştururlar. Komşu
iki hücrenin birbirine bakan yüzeylerindeki iki
düğme desmozom birtakım ince filmentlerle
birbirine bağlı olduklarından hücre içindeki bir
keratin filament ağı bütün bir epitel tabakası
boyunca uzanmış olur.
Hemi desmozomlar düğme desmozomlara benzerler. Aralarındaki fark bunların iki hücreye
değil bir hücreyi bunun alt yüzeyine paralel uzanan bazal laminaya bağlamaktaktır. Bazal
lamina üzerine yerleşen hücreler tarafından sentezlenir. En önemli bileşenleri tip VI kollajen,
proteoglikan, fibronektin ve laminindir.
Sıkı Bağlantı Bölgeleri (tight junctions):
Vücut yüzeyini ve boşlukları kaplayan epitel hücreleri birbirlerine sıkı bir şekilde bağlanarak
bir seçici geçirgenlik seddi meydana getirirler. Böylece bu hücre tabakaları çeşitli
kompartmanları ve bunların içindeki farklı kimyasal yapıdaki sıvıları birbirlerinden ayırırlar.
Epitel hücreleri arasında yer alan sıkı bağlantı bölgeleri, bu hücrelerin birbirlerine
tutunmalarını sağlayarak geçirgenlik seddinin oluşmasında önemli bir rol oynarlar. Bunun en
tipik örneği bağırsaklarda görülür.
Bağırsak epitel hücreleri birbirlerine sıkı bağlantı bölgeleri aracılığıyla bağlanarak bağırsak
lümeni ile kan arasında bir sed oluştururlar. Bu hücreler arasındaki sıkı bağlantı bölgeleri en
10
küçük moleküllerin bile bu seddi geçmelerine izin vermez. Öte yandan vücut için gerekli
maddelerin lümenden kana emilmeleri de gerekmektedir. Bu işi epitel hücrelerinin plazma
membranı içinde yer alan özgül transport proteinleri başarırlar. Bu proteinlerden bazıları
epitel hücrelerinin lümene bakan yüzeyinde yer alırlar ve gerekli maddelerin lümenden hücre
içine geçmelerini sağlarlar.
Diğer bazı transport proteinleriyse bu
maddeleri hücre içinden kana geçirirler.
Bunlar iki şekilde rol oynarlar: ilk olarak
hücrelerin farklı yüzeyindeki membran
transport
proteinlerinin
lateral
difüzyonunu önleyerek bunların yerlerini
korumalarını sağlarlar. İkinci olarak ise
moleküllerin geçişine izin vermeyerek
taşınan
maddelerin
tekrar
lümene
dönmelerini önlerler.
Elektron mikroskop çalışmaları sıkı
bağlantı bölgelerinin yapılarını açıklığa
kavuşturmuştur. Buna göre birtakım
membran içi proteinler hücrelerin
çevresini sarmakta ve bunlar membran
içinde sıralar halinde uzanırlarken bu
sıralar yer yer birbirleriyle bağlantılar
kurmaktadırlar. Sıkı bağlantı bölgeleri iki
komşu
hücrenin
bu
proteinlerinin
birbirleriyle bağ yapmaları sonucu
Geçit Bölgeleri (gap junctions)
Hücreler arasında en yaygın olarak bulunan bağlantı bölgesi tüpü geçit bölgeleridir. Bu
bölgeler iki komşu hücrenin sitoplazmaları arasında bağlantı kurarak bazı moleküllerin bir
hücreden diğerine geçişine olanak sağlarlar. Altı tane connexin proteininin bir araya gelmesi
oluşan connexon birimlerinden olşurlar.
11
Sitoplazma ve Organeller
Hücre zarı ve çekirdek arasında yer alan sitoplazma saydam, jel kıvamında madde ve bu
madde içinde yüzen şekilli unsurlardan oluşmuştur. Sitoplazmayı oluşturan yapılara aşağıda
olduğu gibi değişik isimlendirmeler yapılır:
-Esas plazma (Hyaloplazma): Sitoplazmada hücre organelleri arasındaki sıvıdır. Bu sıvı
bütün hücrelerde bulunur. Protein, su, enzimler, hormonlar, mineraller, yağlar ve
karbonhidratlardan oluşmuştur.
-Metaplazma: Hücrenin fonksiyonu için ortaya çıkan kalıcı fibril şeklindeki yapılar (hücre
iskeleti) metaplazmayı oluşturur.
-Öplazma: Sitoplazmada geçici olarak belli dönemlerde oluşan yapılara verilen isimdir.
Hücre bölünmesi sırasında ortaya çıkıp sonra yok olan mitoz mekiğini, öplazmaya örnek
olarak verebiliriz.
-Paraplazma: Hücrede hayat olayları devamlılık gösterir. Bir kısım maddeler sürekli olarak
yapılır, parçalanır ve canlı maddelerin yapısına katılır. Bazı maddeler ise hücre içinde depo
maddesi gibi bulunurlar.
Hangi yapılar hücrede paraplazmik maddelerdir?
Besin maddeleri, salgı granülleri ve pigmentler hücre içindeki paraplazmik maddelerdir.
Sitoplazmada şekilli unsurlar olarak isimlendirilen yapılardan hücre organellerini,
endoplazmik retikulum, ribozomlar, Golgi kompleksi, lizozomlar, peroksizomlar,
mitokondriyonları sırası ile ayrıntılı olarak inceleyelim.
Endoplazmik Retikulum ve Ribozomlar
Endoplazma retikulum elektron mikroskobu araştırmalarının ortaya çıkardığı organellerden
birisidir. Granüllü ve granülsüz olmak üzere iki tipi bulunur. Granüllü endoplazma retikulumu
membranına düzenli aralıklarla ribonükleoprotein parçacıkları (=ribozom) dizilmiştir.
Ribozomlar tübülüslere tanecikli bir görünüm kazandırırlar.
Ribozomların varlığı granüllü endoplazmik retikuluma bazik boyanma özelliği
kazandırır.
12
Büyük büyülmelerde, endoplazma retikulumu ince tüp şeklinde zarsı yapıların sık sık
dallanmalar göstererek birbirleri ile ağızlaştığı ağ şeklinde düzenlenen bir sistem olduğu
gözlenir. İnce tübüller kimi yerde genişleyerek kese şeklinde görülür. Tübüllerin içi sıvı ile
doludur. Granüllü endoplazma retikulumu hücrenin protein yapımı ile görevli organelidir.
Protein sentezini yapan mikromakineler ribozom tanecikleridir.
Ribozomlar, %65 RNA ve %35 proteinden oluşmuş yuvarlak yapılardır. 120-200 oA
büyüklüğündedirler. Hücrede ya endoplazma retikulumu membranına tutunmuş ya da serbest
tanecikler halinde bulunurlar. Serbest ribozomlar genellikle hücrenin kendi gereksinimi olan
hücre içi proteinleri üretirler. Granülsüz endoplazma retikulumu granüllü tipe oranla daha
ince membranı olan kısa tübülüslerden oluşur. Asidofilik boyanma özelliği gösterirler.
Granülsüz endoplazma retikulumunun görevleri çeşitlidir. Steroid hormon salgılayan
hücrelerde iyi gelişmiştir. Çizgili kaslarda kasılma olayını, kolesterol ve safra yapımını,
ilaçların veya zehirli maddelerin olumsuz etkilerinin yok edilmesi gibi değişik metabolizma
olaylarını düzenler. Bu önemli görevlerini yerine getirebilmek için ise kırktan fazla sayıda
enzim içerirler. Enzimlerin aktiviteleri ırklara, topluluklara ve bireylere göre farklılık gösterir.
Granülsüz endoplazma retikulumun enzim aktivitelerinin bireylere göre farklılığını
nasıl yorumlarsınız?
İlaçlara dayanıklılığın ve yan etkilerin her birey için değişik olmasının nedeni bu şekilde
açıklanmaktadır. Endoplazma retikulum granüllü ve granülsüz şekli aynı hücrede bulunabilir
Bu organel hücrede sentezi yapılan ürünlerin depolandığı ve gidecekleri yönleri gösteren
adresler kazandıkları yer olarak bilinir.
Golgi Kompleksi
Golgi kompleksi, Golgi apparatus ya da Golgi cisimciği olarak da isimlendirilir. Hücrede
çekirdeğin yakınında bulunan yassı keseler ve bunlara eşlik eden veziküllerden oluşmuştur.
Eritrositler ve keratinize epitel hücreleri hariç tüm hücrelerde bulunur. Golgi kompleksinin
boyutları hücre tipine ve salgı aktivitesine göre değişir. Bazı hücrelerde büyük bir tane
bulunuyorken bazı hücrelerde birbirine bağlı çok sayıda (=karaciğer hücresinde en fazla 50
tane) Golgi kompleksi bulunur.
13
Elektron mikroskobi incelemelerinde Golgi kompleksini oluşturan yassı keselerin orta
kısımlarının basık kenarlarının biraz genişlemiş olduğu gözlenir. Yassı keselerin içi sıvı ile
dolu olup düz değil hafif eğilimlidirler. Bu şekilde konveks ve konkav iki farklı yüz ortaya
çıkar. Konveks yüze, giriş yüzü ya da şekillenme yüzü (=cis), konkav yüze ise çıkış yüzü ya
da olgunlaşma yüzü (=trans) ismi verilir.
Granüllü endoplazma retikulumu ve Golgi kompleksi ilişkisi nasıldır?
Granüllü endoplazma retikulumunda sentezlenen ürünler taşıyıcı veziküller aracılığı ile Golgi
kompleksinin şekillenme yüzüne gelir. Belli işlemlerden geçirildikten sonra ise olgunlaşma
yüzüne ulaşarak ürünler cisimciği terkederler. Giriş yüzü endoplazma retikulumu ile
ilişkilidir. Çıkış yüzü ise materyalin iletileceği yöndedir. Proteinler ve lipidler Golgi
kompleksi lümenine giriş yüzünden zar kaynaşması sonucu girerler, daha sonra çıkış yüzüne
doğru ilerleyerek bir olgunlaşma süreci geçirirler. Ürünler modifiye edilerek bir kesecikten
diğerine aktarılır. Çıkış yüzüne ulaşan ürünlerin özel işaretlemeleri yapılmıştır. Hücre içine
bakıldığında en az 10 farklı türde taşıyıcı veziküle rastlanır. Bu veziküllerin üzerinde
moleküler adresler bulunmaktadır. Moleküler adreslere sahip ürün, dış ortama, lizozoma ya da
hücre içine doğru gönderilir. Golgi kompleksinin;
-Endoplazma retikulumda sentezlenen ürünleri tanzim etmek,
-Taşıdığı enzimler nedeni ile sentezlenen ürünlerden bazı çıkarımlar ve eklentiler yapmak,
-Ürünleri konsantre etmek görevleri bulunmaktadır.
Golgi kompleksi bazen şekil değiştirerek yeni bir yapının oluşmasına yol açar. Spermiumun
baş bölgesindeki akrozom adı verilen döllenmeyi kolaylaştıran enzimler bulunduran yapı
Golgi kompleksinin şekil değiştirmesi olayına örnek oluşturur.
Lizozomlar
Sitoplazma içinde yuvarlak şekilli etrafı zarla çevrili organellerdir. Eritrositler dışında her
hücrede bulunurlar. Lizozomlar içinde düşük pH'da etkili 50 eritici enzim tanımlanmıştır.
Enzimler hücrenin kendi sitoplazmasına zarar vermezler. Ölüm sonrası değişimler lizozom
zarının geçirgenliğini bozar. Enzimler sitoplazmayı eritmeye başlar. Histolojik tespit
maddelerinin bu aşamada kullanıldığını görmüştük. Tespit maddeleri lizozom içi enzimleri
makromoleküller halinde oldukları yere çöktürür. Bu şekilde enzimlerin sitoplazmaya
geçişleri engellenir.
Lizozom zarının geçirgenliği bozulursa ne olur?
Hücre kendi kendini yok eder; başka bir deyişle eritir. Lizozomlar hücrenin sindirim organeli
olarak düşünülebilir.
14
Golgi kompleksinin çıkış yüzünden ayrılan içi eritici enzimlerle dolu veziküller homojen
görünümlü primer lizozomlardır. Hücre dışından hücre içine fagositoz ya da pinositoz yoluyla
alınan yapılar (örneğin bakteri ya da besinler) sindirilmek istendiğinde, primer lizozom zarı
ile bu maddelerin çevrelerindeki zar kaynaşır. Lizozomlardaki eritici enzimler keseciğin içine
akar. Sekonder lizozom (=heterofagozom) adı verilen yeni bir yapı gelişir. Eğer hücre içinde
yaşlanmadan dolayı bozulmuş yapılar varsa aynı şekilde primer lizozomlarca sindirilir. Bu
şekildeki sekonder lizozoma otofagozom adı verilir. Otofagozomların sayısı yaşlılıkta, açlıkta
ve doku yaralanmalarında artar.
Peroksizomlar (=Mikrocisimler)
Peroksizomlar sitoplazmada çapları lizozomlara yakın yuvarlak şekilli organellerdir. Katalaz,
peroksidaz ve oksidaz enzimleri içerirler. Oksidazlar yolu ile mitokondriyonlar gibi enerji
üretirler. Ancak bir hücrede enerji üretiminin yaklaşık %90'ı mitokondriyonlarda, %10'u
peroksizomlardadır. Diğer enzimlerin işlevleri çeşitlidir. Peroksizomlar hemen bütün
hücrelerde bulunurlar. Metabolik aktivitesi çok olan hücrelerde sayıca fazladırlar.
Mitokondriyon (=Çoğul ismi mitokondria)
Mitokondriyonlar canlıhücrelerin sitoplazmalarında yavaş hareketleri olan boyları ve
şekillerini değiştiren organellerdir. Zaman zaman bölünebilirler. Genellikle sitoplazma
hacminin %25'ini kapsarlar. Sitoplazma içinde iki kat zarla çevrilidirler. Işık mikroskobu ile
tanecik ya da ipliksi yapılar şeklinde görülürler. Hücre organelleri içinde en büyük olanlarıdır
(0,5-1μ). Enerji üretimi yapan, ayrıca çok sayıda kimyasal işlemin gerçekleştiği organellerdir.
Yiyeceklerdeki karbonhidratlar, aminoasitler ve yağlar mitokondriyon içinde okside edilerek
C02 ve su ile birlikte ATP şeklinde serbest enerji sağlanır. ATP yaşamsal fonksiyonların
hepsinde kullanılan bir enerji tipidir.
15
ATP sentezi için neler gereklidir? Mitokondriyon enzimleri nerede bulunur?
ATP sentezi için birçok enzim gerekir. Bu enzimler mitokondriyon iç ve dış zarı ile
zarlararası kısım ve matrikste bulunur.
Krista nedir?
Mitokondriyon iç zarı Şekil 2.9'da görüldüğü gibi matrikse doğru bir takım çıkıntılar yapar.
Bu çıkıntılara krista adı verilir. Kristaların temel fonksiyonu iç yüzeyi artırmaktır.
Mitokondriyonlar genellikle hücrenin belli bölgelerinde birikirler. Bunun nedeni üretilen
enerjinin tüketileceği bölgeye mümkün olduğunca yakın tutulmak istenmesidir.
Mitokondriyonun çoğalma yeteneği hangi özelliğinden kaynaklanır?
Mitokondriyonlar hakkında önemli buluşlardan birisi kendi DNA molekülünün ve ribozom
gibi organellerinin bulunmasıdır. DNA molekülünün nispeten küçük kopyalarını barındırır.
Bu organelleri yardımı ile bağımsız bölünme ve protein sentez etme özelliklerine sahiptir.
Hücreye yönelik olumsuz koşullardan en çabuk etkilenen organel mitokondriyonlardır.
Hemen büyüyüp şişerler.
ÇEKİRDEK (NUKLEUS) VE ÇEKİRDEKÇİK (NUKLEOLUS)
Çekirdek hücre içi tüm biyokimyasal olayları ve hücre bölünmesini yöneten merkezdir.
Çekirdek katılımla ilgili tüm materyallerin bulunduğu organeldir. Hücre için kesinlikle
gereklidir. Canlı hücrede çekirdek, parlak bir kesecik olarak ayırt edilir.
Çekirdek boyanma özelliği nasıldır?
Hemotoksilin-Eozin gibi asit ve baz boya karışımları ile dokular boyandığında çekirdek mavi
mor renkte görünür. Bütün baz boyalarla çok iyi boyanırlar.
Çekirdeğin şekli genellikle hücre şekli ile uyumludur. Hücrede çoğunlukla orta (sentrik)
yerleşimli olup bir tanedir. Ancak karaciğer hücreleri, testisin erkeklik hormonu (=testesteron)
salgılayan leydig hücreleri gibi hücrelerde çok çekirdek olabilir. Çok çekirdekli hücrelere
polikaryosit denir.
Elektron mikroskobu ile dinlenme durumundaki (interfaz) bir hücrenin çekirdeğinde; çekirdek
zarı (karyoteka), çekirdek sıvısı (=karyoplazma), çekirdekçik (=nukleolus) ve kromatin ağı
bulunur. Çekirdek zarı, hücre zarı gibi çift katlıdır. İki ünit zarın arasında bir açıklık
(=perinükleer aralık) bulunur. İç zar düz yüzlüdür. Dış zara ise ribozomlar tutunmuştur.
Çekirdek zarında yer yer delikçikler (porus) gözlenir. Büyük büyültmelerde delikçiklerin açık
olmadığı ve bir diyafram ile kapalı olduğu gözlenir. Delikçikler çekirdek ve sitoplazma
arasında seçici madde geçişini sağlar.
16
Üstteki şekille ilgili açıklama:
1-Çekirdekçik (Nukleolus) 2-Heterokromatin 3-Ökromatin 4-Granüllü endoplazmik retikulum
5-Çekirdek zarı (=karyoteka) 6-Delikçikler (=Porus) 7- Perinükleer aralık
8-Çekirdek sıvısı (karyoplazma)
Çekirdek sıvısı sitoplazmadan daha yoğun bir sıvıdır. Kromatin ağı ve çekirdekçik arasındaki
boşlukları doldurur.
Çekirdekçik (=nukleolus): Çekirdek içinde sayıları bir veya bir kaç tanedir. Protein
sentezinde görevli ribozomal RNA'nın yapım yeridir. Çekirdekçiğin çevresinde zar bulunmaz.
Çok ve çabuk protein üreten hücrelerde gelişmiş olarak gözlenir. Bazik boyanma
özelliğindedir. Kromatin ağı başlıca DNA (dezoksiribonükleik asit) dan oluşur. DNA
protamin ve histon gibi proteinlerle ince uzun kromatin iplikçiklerini oluşturur. Işık
mikroskobunda kromatin iplikçiklerinin yaptıkları ağ, çok koyu (heterokromatin) ya da çok
açık gözlenebilir (ökromatin). Elektron mikroskobunda kromatin düzenli dağılmış tanecikler
olarak belirir. Ayrıntılı incelemelerde ökromatinin, kromatin tanecikleri seyrek dağıldıkları
için açık renkte, heterokromatinin ise kromatin taneciklerinin bir arada olması nedeniyle koyu
renkle görülmektedir. Metabolizma işlevinin çok hızlı olduğu hücrelerde örneğin sinir
hücrelerinin çekirdeklerinde ökromatin alanları fazla gözlenir.
HÜCRE İSKELETİ
Hücre iskeleti, hücrenin tipik şeklinin korunmasında, hücre hareketlerinin yerine
getirilmesinde ve hücre organellerinin hücre içinde yer değiştirmelerinde görev alır.
Hücre iskeletini hangi yapılar oluşturur?
Mikrotübüller, mikrofilamanlar ve ara filamanlar hücre iskeletini oluştururlar. Ayrı bir başlık
altında topladığımız bu yapıları sitoplazma içinde de anlatmak mümkündür. Hücre iskeleti
unsurları organeller arasında bir çatı oluştururlar. Organellerin çevresini bir ağ gibi kuşatırlar.
Mikrotübüller
17
Mikrotübüller protein yapıda olan, uzun, içi boş silindirik yapılardır. Çapları 24 nm'dir.
Başlıca görevleri hücresel asimetreyi korumak, hücre içi organel iletimine yardımcı olmak,
bölünme olayında kromozom hareketlerini sağlamak ve Golgi kompleksi ile endoplazmik
retikulum arasında materyal iletimini sağlamaktır.
Mikrotübüller bazı kompleks hücre organellerinin yapısına katılırlar. Bunlar hareketli silya
(kinosilya) kamçı (flagella) ve sentriollerdir. Mikrotübüller kinosilya, flagella ve sentriollere
özel bir düzende organize olmuşlardır.
Hücre iskeletinin yapısına katılan elemanlar ile hücre organelleri ilişkisi
1-Mitokondriyon 2-Endoplazmik retikulum 3- Mikrotübüller 4-Ribozom (endoplazmik
retikuluma tutunmuş) 5-Serbest ribozomlar.
Sentrioller bölünmeyen hücrelerde Golgi kompleksi ile çekirdek arasında çift olarak yer alır.
Sentrozom adını alan bu çift yapı 300-500 nm uzunlukta 150 nm çapında olup, silindirik
şekillidir. Sentrozom hücre bölünmesi sırasında mikrotübüllerin oluşturacağı iğ iplikçiklerinin
yerinin belirlenmesinde rol oynar.
Mikrofilamanlar
Mikrofilamanlar aktin molekülünden oluşan ipliksi yapılardır. 5-7 nm çapındadırlar. Hücrenin
fonksiyonu ve şekline göre çok sayıda aktin tipi bulunmaktadır. Aynı hücrede de aktin tipinde
çeşitlilik görülebilir. Mikrofilamanlar hücrede ekzositoz, endositoz, hücre göçü gibi hücre zarı
aktivitelerine katılarak, bölünme sırasında hücrelerin ikiye ayrılmasında bölünme noktasında
yüzük oluştururlar. Kas hücrelerinde kasılmayı sağlarlar ve sitoplazmada organel
hareketlerine eşlik ederler.
Ara Filamanlar
18
Ara filamanlar mikrotübüller ve mikrofilamanlara benzer şekilde olup 8-10 nm çapındadırlar.
Ara filamanları oluşturan proteinler birden fazladır. Örneğin epitel hücrelerinde sitokeratin
bulunurken, düz kas hücrelerinde desmin bulunur. Bu çeşitlilik kanser araştırmalarında
önemlidir. Ara filamanların bulundukları yere göre çeşitli görevleri vardır. Örneğin epitel
hücrelerinde başlıca görevleri mekanik sağlamlılık kazandırmak, su ve ısı kaybını önlemektir.
HÜCRE DÖNGÜSÜ
İnterfaz: Bölünmeyen bir hücre interfaz durumundadır. Bu safhada replikasyon ve bölünme
için hazırlık yapılır.
Çekirdek belirgindir, bir yada daha fazla çekirdekçik ayırt edilebilir.
Kromozomlar kromatin yığını şeklindedir.
G1 Evresi: Ribozomlar ve organellerin miktarı iki katına çıkar
S evresi: Organellerin iki katına çıkarılması devam ederken DNA sentezi olur.
G2 Evresi: Hücrenin mitoza hazırlandığı evredir.
M-Evresi: Mitoz ve sitoplazma bölünmesinin (sitokinez) gerçekleştiği evredir.
Hücre döngüsü, bitkilerde 10-30 saat, hayvanlarda 18-24 saat olmakla birlikte; bazı
organizmalarda 20 dk. gibi kısa ya da günler hatta haftalar sürecek kadar uzun olabilir.
19
Sürelerdeki en büyük farklılık G1 evresinde ortaya çıkar. Örneğin; embriyo hücreleri
çok hızlı bölünür ve G1 yok denecek kadar kısa sürer.
Farklılaşmasını tamamlamış iskelet kası hücreleri ve sinir hücreleri G1 evresinde
tutulu kalmıştır. Ve normal şartlarda asla bölünmezler.
Kalp kası hücreleri ise G2’de tutulu kalmış hücrelerdir. Nedeni bilinmiyor.
** G1’de kalmış bir hücre çekirdeği S fazındaki bir hücreye aktarılırsa, çekirdek etkinleşerek
hücre S fazına girer. Aynı şekilde G2’de kalmış hücre mitozdaki bir hücre ile kaynaştırılırsa
kromozomları hemen yoğunlaşmaya başlar ve kendisi de mitoza girer.
** Hücre döngüsünü kontrol eden maddelere SİKLİN denmektedir. Bunların hücre döngüsü
proteinlerine (CDC) bağlanması yada ayrılmasıyla kontrol sağlanmaktadır.
MİTOZ BÖLÜNME (M!)
PROFAZ
Sentriyoller iki kutba doğru ilerler. Her sentriyol çifti yakınında bir mikrotübül
sistemi ortaya çıkar ve tüm yönlere yayılır. Bu üçlü mikrotübül dizilerine Aster denir.
Sentriyoller kutuplara ilerlerken bazı mikrotübüller aksi yöndeki sentriyollerden
uzanan mikrotübüllere tutunur, böylece polar mikrotübüller oluşturulur. Asterler
polar mikrotübüllerle birlikte sepet benzeri iğler (Mitoz iği) oluşturur. Geç profazda
kromatitlerin sentromeri üzerinde oluşan kinetokor denilen plaklara tutunur.
Çekirdek zarı yavaş yavaş kaybolur.
Krozomlar kısalıp kalınlaşarak belirgin hale gelir. (Kro.lar iki kromatit şeklindedir)
Çekirdekçik kaybolur.
METAFAZ
Metafazdan önce prometafaz denilen kısa bir dönem vardır. Başlangıçta çekirdekte
tamamen rasgele dağılmış olan kro.lar iğ ekvatoruna doğru hareket etmeye başlar. Bu
hareket kinetokor mikrotübüllerini oluşturan tübülin alt birimlerinin büyüyebilme
(birbirine eklenerek) ve kinetokor bağlantılarında sindirilmesinden kaynaklanır.
Prometafaz sonunda her kro. eşit biçimde iki kutup tarafından çekilmiş durumdadır.
Metafaz sırasında kro.lar iğ ortası boyunca dizilirler.
Her ikiz kromatit çiftinin sentromerleri birbirinden ayrıldığı zaman metafaz sona erer.
20
ANAFAZ
İki tam kro. grubu bu evrede birbirinden ayrılır. Tek kromatid kro.ları içeren iki set
farklı kutupları gider.
Kro.lar sentromerlerinden kutuplara çekilirken, zıt kutuplarda bulunan polar
mikrotübüller çapraz bağlantılarla kutupları iterek birbirinden ayırırlar. Bu esnada
mikrotübüllerin boyu uzamaktadır.
Anafaz sonunda hücre birbirinden iyice uzaklaşmış iki kro. grubu içerir.
Sitokinez geç anafazda başlar.
TELOFAZ
Profazın tersidir. Kutuplardaki kro. grubu yeni çekirdek zarları ile çevrilir.
İğ iplikleri kaybolurken kro.lar çözülmeye başlar ve interfaz evresindeki biçimlerine
geri dönerler.
Yeni çekirdeklerin tamamen interfaz haline geçmesiyle telofaz sona erer.
Sitokinez sıklıkla telofaz esnasında tamamlanır.
SİTOKİNEZ
Hayvan hücrelerinde:
Hücreyi saran bir bölünme yarığı oluşumu ile başlar. Bu yarık hücreyi bir uçtan diğer
uca bölene dek derinleşir ve iki yeni hücre meydana getirir.
Aktin-miyozin etkileşmesi sitokinezi gerçekleştirmektedir.
Bitki hücrlerinde:
Hücrenin ekvator bölgesinde hücre plağı denen özel bir zarın oluşumu ile başlar.
Çoğunlukla golgi az miktarda da ER’dan köken alan veziküllerin sitoplazmanın
merkezinde yan yana dizilip sonra bileşirler. Çevresel olarak da yeni oluşmuş
hücrelerin zarlarıyla birleşirler.
Daha sonra orta lamel ve Primer HD öncülleri meydana gelir.
21
MAYOZ BÖLÜNME (R!)
Hücrede kromozom sayısının yarıya indirgenmesi amacıyla yapılan bölünmeye mayoz veya
redüksiyon bölünme denir. Mayoz bölünmenin amacı gerçekte bir çoğalma değil, aksine
eşeysel rekombinasyonları ve bunun sonucunda biyolojik çeşitliliği meydana getirmektir.
Birbirini takibeden iki bölünme şeklinde olan mayozun birinci bölümünde kromozom sayısı
yarıya iner, ikinci bölümünde ise tipik bir mitoz bölünme meydana gelir.
İNTERFAZ I
Mitoz bölünmede olduğu gibidir. Genetik materyal ve organeller kendini eşler.
PROFAZ I
Kromotin iplikler kısalıp, kalınlaşır ve belirgin kromozom şeklini alır. Kromozomlar
üzerindeki kromerler belirgin hale gelir. Bu evreye Leptoten de denir. Çekirdek zarı yavaş
yavaş erir ve sentrozomlar kutuplara doğru hareket eder.Homolog kromozomların her bir
çiftinin yan yana gelip ve birbirinden ayrılmadan kalmalarına zigoten denir. Homolog
kromozomlardaki bu protein eksenler birbirine enine protein köprülerle bağlanarak çok
sağlam synaptonemal kompleksleri oluştururlar. Yapısında 4 kromotid bulunan bu
komplekslere tetrad bu olaya da sinapsis denir. birinci profazın bu evresine ise Zigoten adı
verilir. Sinaps sırasında kromozomlar hem kısalıp hem de kalınlaşırlar. bu evreye de Pakiten
de denir. Bu sırada çok önemli bir olay olan krossing over başlar.
22
METAFAZ I
Mitozda bir çift ikiz kromotid ihtiva eden homolog kromozomlardaki her bir kromozomun
hücrenin ekvator tablasın a hareket etmesine karşılık, mayoz bölünmede ekvator tablasında
yer alan kromozomlar 2 homolog kromozom çifti halindedir ve her birinde ikişer hibrid
kromotid bulunmaktadır.
ANAFAZ I
Bu safhada profazın ortalarında birbirine sinaps yapan homolog kromozomlar birbirinden
ayrılmaya başlarlar.Hibrid kromotidleri ihtiva eden her bir kromozomun iğ ipliklrine takılıp
kutuplara doğru hareket etmesiyle birinci anafaz son bulunur.
TELOFAZ I
Bu safha tümüyle mitoza benzer tek fark ise mitozda yeni teşekkül eden çekirdekte ana
hücreninkine eişt sayıda kromozom bulunmasına karşılık mayoz bölünmede bu kromozom
sayısı yarya inmiştir. Mitoz bölünmedeki her bir kromozom bir kromotid kromozomdur.
Halbuki mayozdakiler bir çift hibrit kromatid ihtiva eden kromozomlar halinde bulunurlar.
MAYOZ II
PROFAZ II
Birinci bölünmenin telofozı ile ikinci bölünmenin arasında bir dinlenme devresi olmadan
çekirdek zarı parçalanır. Birinci iğ iplikçiklerinin doğrultusunu dik yeni iğ iplikçikleri oluşur.
METAFAZ II
Her oğul hücrenin haploıd (n) kromozomu ekvatoral düzlem üzerinde dizilir. Bu evrede
kromozomlar ikili görülür.
ANAFAZ II
Ekvatoral düzlem üzerinde dizilmiş olan diatlar sentromerlerinin uzunlamasına bölünmesiyle
birbirinden ayrılırlar. Her kromatid anafaz kromozomu halinde kutuplara çekilir.
TELOFAZ II
Kromozomların helezonları açılır, dolayısıyla görünmez olurlar. Çekirdek zarları oluşur
sitoplazma bölünür. Böylece bir hücreden 4 tane haploid hücre meydana gelir.
* Sitoplazma bölünmesi
Hayvansal hücrelerde: Çekirdek bölünmesi tamamlandıktan sonra sitoplazma ortadan
boğumlanarak ikiye bölünür ve iki ayrı hücre oluşmuş olur.
Bitkisel hücrelerde: Oluşmuş olan iki çekirdekli hücrenin ortasında bir orta lamel (ara lamel)
oluşur ve hücre duvarına kadar ulaşır. Dolayısıyla birbirine bitişik iki hücre oluşur.
23
Mayoz bölünme sonucunda n kromozomlu 4 hücre (gamet)oluşur.
24