HÜCRELER ARASI
HABERLEŞME VE
İLETİŞİM
HÜCRE HABERLEŞMESİNE GENEL BAKIŞ
• Hücre haberleşmesi canlılığın erken döneminde evrimleşmiştir.
• Yakın zamana kadar tek ve çok hücreli canlılar, hücreler
arası
iletişimin olup olmaması konusunda ayrım gösteriyorlardı.
• Ancak yapılan araştırmalar sonucunda sadece çok hücreli
canlıların değil, tek hücreli canlılarında hücreler arası
haberleşme yaptığı bulunmuştur.
• Bunun sonucu olarak da hücre haberleşme mekanizmasının
öncelikle ilk prokaryotlar ve tek hücreli ökaryotlarda evrimleştiği
daha sonra da bu olayın çok hücreli organizmalara adapte
edildiği düşünülmüştür.
Mayalarda Haberleşme
• Bir fungus olan Saccharomyces cerevisiae eşini kimyasal
haberleşme yöntemi ile bulmaktadır.
• Bu fungusun ‘’a’’ ve ‘’alfa’’ adı verilen iki cinsi vardır. ‘’a
çiftleşme tipi’’ olarak adlandırılan hücreler ‘ ’a faktörü’ adı
verilen kimyasal bir sinyal salgılarlar.
• Bu protein yakındaki bir alfa fungusunun hücre yüzeyindeki
reseptörlerine bağlanır. Aynı şekilde bir alfa fungusuda ‘’alfa
faktörü’’ salgılar ve bu proteinde a fungusunun reseptörlerine
bağlanır.
• Bunun sonucu olarak iki hücre tipi birbirine doğru büyümeye
başlarlar. Sonuçta zıt tipte ki iki hücre içeriği kaynaşır.
Çiftleşme faktörleri alışverişi
Çiftleşme
Yeni a/α hücresi
Cıvık Mantarlarda Hücresel Haberleşme
• Cıvık Mantarlarda buna benzer kimyasal bir uyarıcı
yollarlar. Bu uyarıcının adı ‘’Cyclic AMP (cAMP]’’dır.
• Ortamda besin azlığı veya daha zorlu durumlarda bu
hücreler cAMP salgılarlar ve ortamdaki diğer hücreler bu
sinyale doğru gelerek öbekleşirler.
• cAMP diğer komşu hücrelerin içine girer ve bu şekilde
onların birbirleriyle bir araya gelmesini sağlar. Bu şekilde
bir araya gelen hücreler kalın duvarlı sporlar üretir.
• Sporlar ortam koşulları iyileşene kadar hayatta kalır.
Yerel ve Uzak Mesafeden Haberleşme
• İletişim içinde olan hücreler, birbirlerine yakın veya uzak olma
durumuna göre farklı haberleşme tipleri kullanırlar.
•
• Eğer hücreler birbirlerine yakınsa o zaman ‘’LOKAL(YEREL)
HABERLEŞME’’ dediğimiz durum ortaya çıkar.
•
• Bir hayvan hücresi yakınında ki başka bir hücreye sinyal yollayarak
(büyüme faktörleri) onun büyüme ve çoğalmasını aktive eder. Bu
şekilde görülen lokal haberleşme tipi ‘’Parakrin Haberleşme’’ olarak
adlandırılır. Parakrin haberleşmede sinyali veren ve sinyali alan hücre
tipi aynı değildir.
• T lenfositler antijenik uyarıya karşı ‘’Otokrin Haberleşme’’yi
kullanırlar. Bu sayede T lenfosit sayısı artar ve bağışıklık antijene
karşı güçlenmiş olur.
• Otokrin Haberleşmede sinyali veren ve alan hücreler aynı tip
hücrelerdir. Anormal otokrin haberleşme kanser hücrelerinin
kontrolsüz şekilde çoğalmasına neden olur.
Yerel ve Uzak Mesafeden Haberleşme
• Lokal Haberleşme gösteren hücrelerdeki bir diğer haberleşme tipi
‘’Sinaptik Haberleşme’’dir.
• Bu özelleşmiş haberleşme tipi hayvanlardaki sinir sisteminde
gerçekleşir. Nöron, kimyasal bir haberci olan ‘’Nörotransmitter madde’’yi
oluşturur. Çevreden gelen uyarıcı etkenler sonucunda nöronun aksonu
boyunca ilerleyen elektriksel ileti, akson ucunda bulunan vezikülleri
harekete geçirir.
• Harekete geçen bu veziküller iki nöron arasındaki sinaptik boşluğa
nörotansmitter maddeyi salar.
• Bu madde hedef nöron zarında ki reseptörlere tutunur ve iletişim bu
şekilde gerçekleştirilir.
• Sinaptik haberleşmede iletim çok hızlıdır. (Saniyede 100 metre hız gibi)
Parakrin haberleşme
Uzak mesafeden haberleşme
Sinaptik haberleşme
Yerel ve Uzak Mesafeden Haberleşme
• Lokal haberleşme yüksek organizmalı canlılarda hücrelerin
davranışlarını tek başına koordine etmekte yetersizdir.
• Daha uzak bölgelere haberleşmeyi sağlamak için hayvanlarda
‘’endokrin hücre’’ adını verdiğimiz özelleşmiş sinyal hücreleri
bulunur. Bu hücreler hormon adı verilen kimyasalları
kullanırlar.
• Hormonlar kan dolaşımı yoluyla, uzaktaki hedef hücrelere
ulaşırlar.
• Hipofiz, tiroit, paratiroit, pankreas, adrenal bezler ve gonatları
içeren endokrin bezler sayesinde 50’den fazla farklı hormon
üretilir.
Haberleşme
• Bu haberleşme yolları dışında hücreler doğrudan temas
yoluylada haberleşebilirler.
• Hayvan hücreleri diğer hücrelerle temasta kalmak için çeşitli
bağlantı yolları barındırırlar.
• Bu bağlantılar doğrudan komşu hücre sitoplazmalarını birbirine
bağlar.
• Böylece sinyal gönderen hücre bileşiği, komşu hücre
sitoplazmaları arasında serbestçe hareket edebilir.
• Ayrıca hücre yüzeylerindeki moleküller aracılığıyla birbirleriyle
doğrudan temas kurabilirler.
Hücre
bağlantıları
Hücrelerin birbirini tanıması
HÜCRE İLETİŞİMİNİN AŞAMALARI
1) Sinyal Alma: Hedef hücrenin hücre dışından gelen
sinyali algılaması aşamasıdır. Kimyasal bir sinyal, hücre
yüzeyinde bulunan reseptör bir proteine bağlandığı
zaman algılanır.
2)
Sinyal Aktarımı: Sinyal molekülü, bağlandığı reseptör
proteini değişikliğe uğratır ve sinyal aktarım sürecini
başlatır. Bu süreçte sinyal, hücrenin algılayabileceği bir
forma dönüştürülür.
3) Cevap: Bu aşamada algılanan sinyal, o sinyale özgül bir
hücresel cevabı tetikler.
SİNYAL ALMA VE SİNYAL AKTARIMININ
BAŞLAMASI
• Belirli bir kimyasal sinyalin hedefi olan hücre, bu sinyal
molekülünü tanıyan reseptör proteinlere sahiptir.
• Reseptör protein, hedef hücrenin kimlik kartı gibidir.
• Örneğin; epinefrin kan içerisinde dolaşırken pek çok hücre
tipiyle karşılaşmasına rağmen sadece belirli hücreler bu
hormonu algılar ve ona cevap verirler.
• Haberci molekül bir reseptör proteine bağlandığı zaman onun
biçim değiştirmesine neden olur.
• Biçim değiştiren bu reseptör protein diğer hücresel moleküllerle
etkileşime açık hale gelir.
Hücre haberleşmesi
SİNYAL ALMA VE SİNYAL AKTARIMININ
BAŞLAMASI
• Sinyal reseptörlerinin bir çoğu plazma zarındaki proteinlerdir.
• Kısaca açıklamak gerekirse gelen sinyal molekülü suda
çözündüğü ve plazma zarından geçemeyecek kadar büyük
olduğu için kendini hücrenin içine aktaracak bir taşıyıcıya
ihtiyaç duyar. Bu taşıyıcıda plazma zarındaki reseptör
proteindir. Reseptör protein sinyal molekülün bağlanmasıyla
şekil değişikliğine uğrar ve sinyali hücre içine yollar.
• 3 temel zar reseptör tipi vardır.
1. G-Proteine bağlı reseptörler,
2. Tirozin-kinaz reseptörleri ve
3. İyon kanalı reseptörleridir.
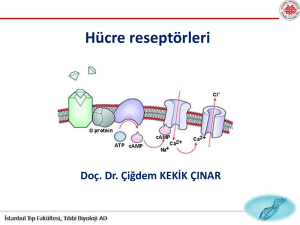
1) G- Proteine Bağlı Reseptörler (GPCR):
• Bu reseptör ‘’G protein’’ adı verilen bir protein yardımıyla iş görür.
• Mayalardaki çiftleşme faktörü, epinefrin ve diğer hormonlar
ile nörotransmitter madde gibi sinyal molekülleri G-Proteine
bağlı reseptörleri kullanırlar.
• Bu reseptör, plazma zarını kateden yedi tane alfa heliks içerir.
G- Proteine Bağlı Reseptörler:
• G Proteini zarın sitoplazmik kısmındadır. Üzerinde GDP veya GTP
bulundurabilir.
• Proteinde GDP molekülü var iken sistem inaktif, üzerinde GTP var
iken ise sistem aktiftir.
• Sinyal molekülü reseptöre gelir ve ona bağlanır. Daha sonra
reseptörde biçim değişikliği meydana gelir ve bu sayede reseptör G
proteinine bağlanır. G Proteinine bağlandığı zaman proteindeki GDP,
GTP ile yer değiştirir. Sonunda protein aktif hale geçer.
• Aktif hale geçen protein, genellikle bir enzim olan başka bir proteine
bağlanarak onun aktivitesini değiştirir ve hücresel bir cevap
oluşturulmasını sağlar.
• Enzim ve G Proteinindeki değişiklikler geçicidir. Çünkü G Proteini aynı
zamanda GTPaz gibi davranır ve işlem tamamlanınca GTP’yi
hidrolizleyerek tekrar GDP’ye çevirir ve inaktif forma döner.
G- Proteine Bağlı Reseptörler:
G- Proteine Bağlı Reseptörler:
• G- Protein Reseptör sistemleri son derece yaygın olup çeşitli
işlevler gerçekleştirirler.
• Embriyonik gelişimde son derece önemlidir.
• Örneğin; belirli bir G-Proteinden yoksun fare embriyoları
normal kan damarlarını geliştiremez ve uterus içinde ölürler.
• Aynı zamanda duyu algılamasında da iş görürler. İnsanlarda
ki görme, tat alma ve koklama bu tip proteinlere bağlıdır.
• Kolera ve boğmaca etmeni olan bakteriler, G-Protein işlevini
aksatan toksinler üreterek kişiyi hastalandırırlar.
2) Tirozin-Kinaz Reseptörleri (RTK):
• Temel reseptör sınıflarından birisidir. Reseptörün sitoplazma tarafında
kalan kısmı Tirozin-kinaz adı verilen bir enzim taşır. Bu enzim,
Reseptör protein üzerinde bulunan Tirozin aminoasidine
ATP’den fosfat grubu aktarımını sağlar.
• Tirozin-kinaz reseptörleri, sinyal molekülleri gelmeden önce bağımsız
polipeptidler şeklindedir.
• Bu polipeptid yapının ucunda sinyali almaya yarayan bir reseptör, zarı
kateden bir alfa helix yapı ve sitoplazma tarafında bulunan tirozin
aminoasidlerinden oluşur.
• Sinyal molekülü reseptöre ulaştığı zaman onun konfigürasyonunu
değiştirmez bunun yerine iki polipeptid zincirin bir araya gelmesini
sağlar. Bunun sonucunda her bir polipeptid zinciri, diğerinde bulunan
tirozin aminoasitlerine fosfat ekler.
Tirozin-Kinaz Reseptörleri:
• Özetle G- Protein reseptöründen farklı olarak bunlarda şekil değişikliği
yerine bir araya gelme (agregasyon) ve reseptördeki tirozinlerin
fosforlanması görülür.
• Bu şekilde aktive olan kompleks, hücre içerisindeki aktarım proteinleri
tarafından tanınır.
• Aktarım proteinleri özgül tirozinlere bağlanır ve kendileride aktif hale
geçerler.
• Bu sayede aktive olan aktarım proteinleri özgül birçok hücresel
cevaba neden olan sinyal aktarım yollları başlatır.
• G-Proteine bağlı reseptörlerle en büyük farkı; Tek bir sinyal
molekülünün Tirozin-Kinaz reseptörleri sayesinde çok sayıda yolu
tetikleyebilmesidir.
3) İyon Kanalı Reseptörleri:
• Bu reseptörler, sinyal molekül varlığında açılıp kapanan
kanallar şeklindedir.
• Bu kanalların açılması sayesinde Na ve Ca gibi iyonların
geçişine izin verilir ya da bunların geçişi engellenir.
• Sinyal molekülü, kanal proteininin hücre dışındaki özgül bir
kısmına bağlanır.
• Kanal proteinde ortaya çıkan şekil değişikliği sonucu, bir
iyonun hücre içi derişimi değişir.
• Sinyal molekül varlığında açılıp kapanan iyon kanalı sistemi
sinir hücreleri üzerinde çok etkilidir.
Hücre içindeki reseptörler;
• Hedef hücrelerin stoplazması yada çekirdeğinde bulunan
pr.lerdir. Kimyasal haberci buraya ulaşabilmek için plazma
zarından geçebilmelidir.
• Bazı önemli sinyal molekülleri ya hidrofobik yada hidrofobik
kısmı geçebilecek kadar küçük olduğundan başarılı olabilir.
• Örn; hayvanlardaki steroid hormonlar ve tiroid hormonları
• Hücre içinde reseptöre sahip başka kimyasal mesajcı; Nitrik
oksit (NO) gazının küçük molekülleri de zar fosfolipidlerinden
kolayca geçebilir.
Hücre içindeki reseptör ile steroid hormon arasındaki
etkileşim
II. SİNYAL AKTARIMI
• Moleküler etkileşim şellaleleri sinyalleri reseptörlerden
hücre içindeki hedef moleküllere ulaştırırlar.
• Sinyal molekülü fiziksel olarak aktarılmayıp bilginin
aktarımı şeklindedir.
• Aktarım yolundaki her basamakta sinyal farklı bir forma
aktarılır ve bu form genellikle proteindeki konformasyonel
değişikliklerdir.
• Konformasyonel değişiklik ise FOSFORİLASYON aracılığı
ile olur.
PROTEİN FOSFORİLASYONU
• Hücre zarındaki bir reseptöre, ona özgü bir sinyal molekülün
bağlanması, sinyal aktarım yolundaki ilk basamağı tetikler ve
hücrenin belirli bir yanıt vermesine yol açar.
• Sinyal ile aktive olmuş bir reseptör, diğer bir molekülü aktive
eder.
• Bu süreç hücresel yanıtı oluşturacak molekül aktive olana
kadar devam eder.
• Aktarım molekülleri denen bu moleküllerin çoğu proteindir ve
hücre iletişiminin temel mekanizması proteinlerin etkileşimine
dayanır.
PROTEİN FOSFORİLASYONU
• Protein fosforilasyonu sayesinde, proteinde konformasyonel
değişiklikler meydana gelir. Protein fosforilasyonu, protein aktivitesini
denetlemek için hücre tarafından kullanılan çok yaygın bir
mekanizmadır.
• Protein kinaz enzimleri sayesinde bu protein yapılara, ATP’den fosfat
grupları eklenir. Protein fosfatazlar defosforilasyonda görev yapar.
• Bu şekilde devam eden fosforilasyon olayından sonra ‘’Fosforilasyon
Şelaleri’’ dediğimiz olay oluşur. Fosforilasyon şelalesi sayesinde
sinyalin çoğaltılması sağlanır.
• Sinyalin çoğaltılması sonucunda, örneğin karaciğer ya da kas
hücresinin yüzeyindeki reseptörlere bağlanmış olan az sayıdaki
epinefrin molekülü, glikojenden yüz milyonlarca glukoz molekülü
oluşturulmasına yol açar.
İKİNCİ MESAJCILAR (HABERCİLER)
• Sinyal aktarım yollarındaki moleküllerin tümü protein değildir;
bunlar suda çözünebilen, küçük moleküller ya da iyonlar da
olabilir. Bu moleküllere ikinci mesajcılar adı verilir.
• İkinci mesajcılar hem küçük, hem de suda çözünebilir
olduklarından, tüm hücreye difüzyon yoluyla kolayca dağılır.
• Örneğin cAMP adı verilen ve epinefrin tarafından başlatılan
sinyali karaciğer ya da kas hücresinin zarından hücre içine
taşıyan molekül bir ikinci mesajcıdır. cAMP karaciğer ya da kas
hücresinde glikojen yıkımını başlatır.
• En yaygın olarak kullanılan ikinci mesajcılar cAMP ve kalsiyum
iyonlarıdır.
cAMP’nin ikincil mesajcıl olarak iş görmesi
Ca +2 iyonları ve İnozitol Trifosfat
• Sitozolik Ca
derişimini
artırarak sinyal aktarımı
• Hayvan hücrelerinde kas
kasılması, belirli bileşiklerin
salınması ve hücre
bölünmesi gibi cevaplara
neden olur.
• Hücreler hem G-pr. hemde
tirozin kinaz yollarında Ca +2
‘u ikinci haberci olarak
kullanırlar.
• Kalsiyum salınmasına yol
açan yollar, DAG
(diaçilgliserol) ve IP3 (inozitol
trifosfat) gibi ikincil
mesajcıları da içerir.
+2
diaçil gliserol
Haberleşme yollarında kalsiyum ve inozitol trifosfatın
görevleri
III. SİNYALLERE VERİLEN HÜCRESEL YANITLAR
(Hücresel aktivite düzeni)
• Sinyale verilen yanıt akla gelebilecek her çeşit hücresel
etkinlik olabilir. Metabolizma, hücre iskeletinin yeniden
düzenlenişi, genlerin aktivasyonu, hücre göçü, hücrenin
konumlanması, hücre bölünmesi bu etkinlikler arasında
sayılabilir.
• Hücrenin bir sinyale verdiği cevap, sitoplazmadaki
etkinliklerin düzenlenmesi ya da nukleusta
transkripsiyon yapılması olabilir.
Sinyale verilen
ç e k i r d e k
cevabı=gen
aktivitesinin
düzenlenmesi
Bir büyüme faktörü
tarafından özgül bir
genin aktivasyonu
Sinyale verilen sitoplazmik cevap
III. SİNYALLERE VERİLEN HÜCRESEL
YANITLAR
• Farklı hücreler aynı hücre dışı sinyal molekülüne farklı şekilde
yanıt verebilir.
• Örneğin, asetilkolin nöroaktarıcısı iskelet kası hücrelerinin
kasılmasını tetiklerken, kalp kası hücrelerinin kasılma kuvveti
ve hızını düşürür.
• Bu, çizgili kastaki asetilkolin reseptör proteinlerinin kalp
kasındakilerden farklı olmasına bağlıdır.
• Ancak reseptör farklılıkları her zaman farklı etkileri açıklamaya
yetmez. Çoğu zaman, aynı sinyal molekülü tıpatıp aynı
reseptör proteinine bağlansa da farklı hedef hücrelerde
bambaşka yanıtlara yol açar; bunun nedeni ise hücrenin gelen
sinyalleri birleştirip yorumlayacağı hücre içi düzeneğin farklı
olmasıdır.
SİNYALİN SONLANDIRILMASI
•
Bir hücrenin hücre dışı bir sinyale normal bir yanıt vermesi için,
başlangıçtaki sinyali ortadan kaldırıp sinyal aktarım yolunu kapatacak
mekanizmalara sahip olması gerekir.
• Protein kinazların etkileri hücredeki protein fosfatazlar tarafından
durdurulur.
• Protein fosfatazlar proteinlerdeki fosfat gruplarını uzaklaştırır. Bu sayede
mekanizma tersine dönmüş olur.
• Fosfodiesterazlar, cAMP’ı AMP’ye çevirerek sinyali durdurabilir.
• G protein ile eşleşmiş yollarda bir G proteinin GTPaz kısmı GTP’yi GDP’ye
çevirerek G proteinini inaktif eder.
• Sinyal-reseptör sisteminin endositozla hücre içine alınması veya olduğu
yerde inaktive edilmesiyle de sinyal sonlandırılır.