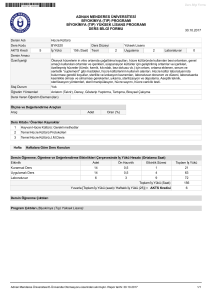
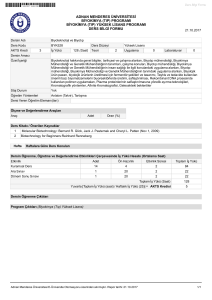
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Biyokimya Laboratuvarı I – Güz
0
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
İçindekiler
Laboratuvar Kuralları ve Güvenlik……………………………………….....2
Deney I
Amino Asitlerin Titrasyon Eğrileri Ve İzoelektrik Nokta Tayini ……………….…....3
Deney II
Amino Asitlerin Kağıt Kromatografisi İle Ayrılmaları………………………….......7
Deney III
Amino Asitlerin Ve Proteinlerin Bazı Özelliklerinin İncelenmesi .…………………11
Deney IV
Tampon Çözeltiler…………………………………………………..……..…………….19
Deney V
Katekolün Oksidasyonuna Polifenol Oksidaz Enziminin Etkisi………………………...23
Deney VI
Proteinlerin Saflaştırılması Ve Karakterizasyonu…………………………………….....32
Deney VII
Proteinlerin Kantitatif Tayini………………………………………………...………….39
Deney VIII
Proteinlerin Elektroforez İle Ayrılmaları Ve Molekül Ağırlıklarının Tayini………....…45
Deney IX
Proteinlerin N- Terminal Amino Asitlerinin İnce Tabaka Kromatografisi İle Belirlenmesi……54
Deney X
Memeli Dokularından DNA İzolasyonu Ve Bazı Özelliklerinin İncelenmesi………..…60
Kapak resmi: İnsan insülin parçalayıcı enziminin beta-amiloid (mavi) ile verdiği
kompleksin yapısı
Biyokimya Laboratuvarı I – Güz
1
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Laboratuvar Kuralları ve Güvenlik
Biyokimya laboratuvarı güz döneminde, protein saflaştırılması ve karakterizasyonu,
enzim kinetiği ve DNA izolasyonu gibi biyokimyada yaygın olarak kullanılan tekniklerle
ilgili genel prosedürlerin tanıtılması hedeflenmektedir. Ayrıca biyokimyada sık olarak
kullanılan bazı cihazların kullanımı öğretilecektir.
Her labarotuvardan önce o gün yapılacak olan deneyi mutlaka dikkatle okuyunuz. Her
deneyden önce bir kısa sınav yapılacaktır. Bu sınav sonucunda sıfır almış olan öğrenci o
deneye alınmaz.
Laboratuvarda çalışırken laboratuvar önlüğü ve (steril olmayan) laboratuvar eldiveni
kullanılır. Önlük ve eldivenlerinizi laboratuvardan ayrılıncaya kadar çıkarmayınız. Bazı
yumuşak lensler laboratuvar kimyasalları ile tepkimeye girerek rahatsızlık
oluşturacağından, eğer mümkünse kontak lenslerinizi laboratuvara girmeden önce
çıkarınız. Deney esnasında asistanların talimatlarını dikkatle uygulayınız. Deney
bitiminde tüm cihazları kapatıp, çalıştığınız yerin temizliğini iyi bir şekilde yapınız.
Bazı kimyasallar zehirli, mutajenik, karsinojenik veya teratojenikdir (doğum kusurlarına
yol açan). Bu kimyasalların kullanımında ve atılmalarında laboratuvardaki görevlilerin
yaptığı uyarıları dikkate alınız.
Tüm öğrencilerin bir laboratuvar defteri tutmaları istenmektedir. Deney esnasında
yaptığınız her bir ölçümü, deney aşamasını, dikkatle laboratuvar defterinize not alınız.
Her deneyin tamamlanmasından sonra en geç bir sonraki deneyden bir gün önce deney
raporlarınızı ilgili asistana teslim ediniz. Bu raporlar aşağıdaki şekilde hazırlanmalıdır.
Başlık: Deneyin adı
Materyal ve Metod: Kısaca deneyin yapılışı ve kullanılan kimyasallar
Sonuçlar ve tartışma: Deney sonunda elde ettiğiniz veriler ve yorumlarınız
Çalışma Soruları: Çalışma sorularına verdiğiniz yanıtlar
Öğrenciler deneyleri gruplar halinde yapmaktadırlar ancak her bir öğrenci kendi
yorumlarını, kendi hazırladığı deney raporu ile sunmalıdır. Birbirinin aynı olan raporlar
kabul edilmez. Aynı şekilde çalışma sorularına cevap verilmemiş deney raporları da
kabul edilmez. Deneylerin değerlendirilmesinde en önem verilen kısım tartışma kısmıdır.
Mutlaka her bir deneyin neden gerçekleştiği (veya neden gerçekleşmediği) konusunda
kendi açıklamalarınızı ekleyiniz.
Her dönemde bir ara sınav yapılır. Laboratuvar kısa sınavlarının %10’u deney
raporlarının %20’si ve ara sınavın %70’i alınarak öğrencinin vize notu saptanır. Ara
sınavdan sonra yapılan deneyler için de laboratuvar kısa sınavlarının %10’u deney
raporlarının %20’si ve finalin %70’i alınarak öğrencinin final notu saptanır.
Biyokimya laboratuvarında devam zorunluluğu %80’dir. 3 kez laboratuvara gelmeyen bir
öğrenci devamsızlıktan kalmış olur ve sınavlara alınmaz.
Biyokimya Laboratuvarı I – Güz
2
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY I
AMİNO ASİTLERİN TİTRASYON
İZOELEKTRİK NOKTA TAYİNİ
EĞRİLERİ
VE
Amfoterik maddeler olan amino asitlerde iki fonksiyonel grup bulunur. Bunlar
karboksilik asit (-COOH) ve amino (-NH2) gruplarıdır. Bu fonksiyonel gruplar nötral
ortamda iç tuz oluştururlar.
Nötral Ortamda:
(İç Tuz)
Asidik Ortamda:
(Katyon)
Bazik Ortamda:
(Anyon)
Yukarıdaki denklemlerden de görülebileceği gibi amino asitlerin amino grubu hidrojen
iyonu kabul edebilir, karboksil grubu ise hidrojen verebilir. Bu sebepten dolayı amino
asitler hem asitler hem de bazlarla titre edilebilirler, yani amfoterik özellik gösterirler.
Amino asitlerde titre edilebilir en az iki grubun olması, en az iki denge sabiti (K) ve iki
iyonizasyon sabiti (Ki) olduğu anlamına gelir. İzoelektrik nokta (pI), aminoasitte
bulunan artı ve eksi yüklerin birbirine eşit olduğu, yani amino asidin net yükünün 0
olduğu pH değeridir, yani K1 ve K2’nin dengede olduğu noktadır.
Biyokimya Laboratuvarı I – Güz
3
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Asit-baz titrasyonları protonların kademeli olarak eklenmesi veya uzaklaştırılmasıdır.
Aşağıdaki şekilde glisinin titrasyon eğrisi gösterilmiştir.
Şekilden glisinin iki tamponlama aralığına
sahip olduğu görülmektedir. Bunlar pH
2.34 ve pH 9.6 merkezli +/-1 pH aralığına
sahip
dikdörtgen
alanlar
olarak
gösterilmişlerdir.
Glisin
bu
pH
aralıklarında tampon çözelti olarak
kullanılabilir. Şekilden de görüldüğü gibi
glisinin izoelektrik noktasının teorik değeri
5.97’dir.
İyonlaşabilen yan zincire sahip amino
asitlerde ise üç bölgeli bir titrasyon grafiği
beklenmelidir.
Şekil 1.1- Glisinin titrasyon eğrisi
KÂĞIT
Şekil 1.2- Glutamik asidin titrasyon eğrisi
Biyokimya Laboratuvarı I – Güz
Şekil 1.3- Histidinin titrasyon eğrisi
4
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
KİMYASALLAR
HCl, NaOH, Glisin.
YÖNTEM
50 mL 0.1 N Glisin çözeltisi hazırlanır. pH metrenin kalibrasyonu yapıldıktan sonra,
hazırlanan çözeltiden 20 mL’lik bir hacim bir behere aktarılarak pH metre yardımı ile pH
tayin edilir. Daha sonra 0.1N HCl ile titre edilir. Her 1 mL asit ilavesinden sonra beher
karıştırılarak pH ölçümü yapılır. İşleme pH göreceli olarak sabit kalıncaya kadar devam
edilir. pH metrenin elektrodu destile su ile yıkanır, kurulanır.
Hazırlanmış olan 0.1N Glisin çözeltisinden bir başka behere 20 mL daha alınır ve pH
metre yardımı ile pH ölçümü yapılır. Daha sonra 0.1N NaOH ile titre edilir. Her 1mL baz
ilavesinden sonra beher karıştırılarak pH ölçümü yapılır. İşleme pH göreceli olarak sabit
kalıncaya kadar devam edilir.
Elde edilen değerler veri tablosuna işlenir.
Asit titrasyonu veri tablosu:
Eklenen
HCl (mL)
0
1
pH
Eklenen
HCl (mL)
pH
Eklenen
HCl (mL)
pH
Eklenen
HCl (mL)
pH
Baz titrasyonu veri tablosu:
Eklenen Baz
(mL)
0
1
pH
Eklenen Baz
(mL)
pH
Eklenen Baz
(mL)
pH
Eklenen Baz
(mL)
pH
Biyokimya Laboratuvarı I – Güz
5
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
VERİLERİN KULLANIMI
Milimetrik grafik kağıdı kullanarak x eksenine eklenen asit ve eklenen baz hacmini, y
eksenine ise ölçülen pH değerlerini koyarak grafiği çiziniz ve grafikten pKa ve pKb
değerlerini ve pI değerini bulunuz. Grafik üzerinden elde ettiğiniz verileri teorik değerler
ile karşılaştırınız.
Şekil 1.4- Glisinin asit ve bazla titrasyon eğrisi
ÇALIŞMA SORULARI
1- Glutamik asit ve histidinin pI değeri nasıl hesaplanır? Aminoasitlerin farklı pI
değerlerine sahip olmalarının önemi nedir?
2- Genel amino asit denklemi kullanılarak izoelektrik noktası formülü nasıl çıkarılır?
Biyokimya Laboratuvarı I – Güz
6
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY II
AMİNO ASİTLERİN KÂĞIT KROMATOGRAFİSİ İLE
AYRILMALARI
Kromatografi bir karışımdaki maddelerin iki faz arasında farklı dağılması esasına
dayanan bir ayırma yöntemidir. Sistemdeki fazlardan biri sabit (stasyoner) faz, diğeri ise
hareketli (mobil) faz olarak adlandırılır. Sabit faz genellikle destek ortamı oluşturmak
amacı ile kullanılırken, hareketli faz, karışımı oluşturan bileşenleri taşımak amacı ile
kullanılır. Karışımı oluşturan bileşenler hareketli ve sabit faz ile farklı oranlarda
etkileşerek farklı hızlarda yürürler. Bu esasa dayalı olarak ayırma yapan yöntemlere
kromatografik yöntemler denir ve ayırmayı sağlayan kuvvete göre adsorpsiyon, dağılım,
iyon değişim, jel geçirgenlik ve afinite (ilgi) olmak üzere beş sınıfa ayrılır.
Kromatografik yöntemler biyokimya alanında, istenen bir maddenin saflaştırılması
(örneğin bir bitki ya da bir hayvan dokusundan proteinlerin elde edilmesi), seyreltik
çözeltilerin konsantre edilmesi, bir maddenin saflığının kontrol edilmesi ve
organizmadaki metabolitlerinin belirlenmesi gibi alanlarda kullanılır.
Kâğıt kromatografisi, dağılım kromatografisinin özel bir şeklidir. Burada kullanılan
Whatman kromatografi kâğıtları saf selülozdan, herhangi bir katkı maddesi
kullanılmadan üretilmiştir ve düzgün gözenek boyutuna sahiptir. Whatman kâğıdı,
durağan faz olarak adsorplanmış su içerir. Hareketli fazın kapiler etkisiyle ayrımı
yapılacak olan maddeler Whatman kâğıdı boyunca hareketli faz ile etkileşimlerine göre
farklı hızlarda ilerlerler.
Karışımı oluşturan bileşenler renkli ise kâğıt üzerinde yürüdükleri yerlerde yuvarlağa
yakın lekeler şeklinde görülürler. Eğer renksiz iseler üzerlerine bir reaktif püskürtülerek
ya da UV ışığı altında renk göstermeleri ile görünür hale getirilirler. Leke büyüklüğü ve
renk şiddeti kullanılarak kantitatif veya semi-kantitatif analiz yapılabilir.
Retensiyon Faktörü (Rf) lekelerin merkez noktasının başlangıç noktasına uzaklığının,
çözücü sınırının başlangıç noktasına olan uzaklığına bölünmesi ile elde edilir. Rf değeri
bir karışımı oluşturan bileşenlerin tanımlanması için kullanılır. Aynı şartlarda (sabit ve
hareketli fazın cinsi, sıcaklık vb.) kromatografisi yapılan her bir maddenin karakteristik
bir Rf değeri vardır.
Biyokimya Laboratuvarı I – Güz
7
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 2.1- Amino asitlerin ninhidrin reaksiyonu mekanizması
KİMYASALLAR
Asetik asit, n-butanol, ninhidrin, aspartik asit, lösin, valin, arjinin, prolin, fenilalanin,
HCl, Whatman No.1 kromatografi kâğıdı.
YÖNTEM
n-butanol: asetik asit: su (65:15:25) karışımı hareketli faz olarak hazırlanır. Kromatografi
tankına 10 mL (yaklaşık 1 cm yüksekliğinde olacak şekilde) çözücü konulur ve tankın
ağzı kapatılır. Tankın çözücü buharı ile dengeye gelmesi için kromatografi kâğıdı
yerleştirilinceye kadar kapağı açılmaz.
Biyokimya Laboratuvarı I – Güz
8
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Size verilen 5cm x 8cm boyutundaki Whatman No:1 Kromatografi kağıdının kısa
kenarından 1.2 cm uzaklığında kurşun kalemle bastırmadan çok hafif bir çizgi çizilir ve
bu çizgi üzerinde 1cm aralıkla 4 nokta işaretlenir.
1N HCl içinde çözünmüş aspartik asit, lösin, valin,
arjinin, prolin, fenilalanin çözeltilerinden grubunuza
verilmiş olan üçü ve amino asit karışımı bilinmeyen
örnek, ayrı ayrı işaretlenmiş 4 noktaya temiz bir
kapiler tüp yardımı ile tatbik edilir. Tatbik noktasının
çapı 4 mm’yi geçmemelidir. Birden fazla damla
tatbikinde, her damla tatbikinden sonra leke yerinin
kurutulması ve ikinci damlanın ondan sonra tatbik
edilmesi gereklidir.
Tatbik edilen noktalar kurutulduktan sonra kâğıt, bir
pens ile tutularak tatbik noktaları aşağıya gelecek
şekilde tanka yerleştirilir. Kâğıdın, tankın
kenarlarına değmemesine ve tatbik noktalarının
çözücü ile doğrudan temas etmemesine dikkat edilir.
Çözücünün ilerlemesi takip edilir ve üstten yaklaşık
Şekil 2.1- Kromatografi kağıdı
0.5 cm boşluk kalıncaya kadar yürütülür (Bu işlem yaklaşık 45 dakika sürer). Çözücünün
yürüdüğü mesafe hemen kurşun kalemle işaretlenerek kâğıt etüvde kurutulur.
Etüvden çıkartılan kâğıdın üzerine çeker ocakta ninhidrin reaktifi püskürtülür ve
kurutulmak üzere yeniden etüve konulur. Oluşan mor lekelerin çevresi kurşun kalemle
işaretlenerek Rf değerleri hesaplanır ve veri tablosuna kaydedilir. Karışımda bulunan
amino asitlerin hangileri olduğu hesaplanan Rf değerleri ile karşılaştırılarak tayin edilir.
Amino Asit
Lekenin Rengi
Rf Değeri
Aspartik Asit
Lösin
Valin
Arjinin
Prolin
Fenilalanin
Örnek
Şekil 2.2- Yürümesi tamamlanmış olan kromatografi kağıdı.
Biyokimya Laboratuvarı I – Güz
9
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
ÇALIŞMA SORULARI
1- Rf değerlerini hesapladığınız amino asitlerin yürüme hızlarının farklı olmasının
nedenlerini araştırınız. Bu amino asitlerin formülleri aşağıda yer almaktadır.
2- Glutamik asit, histidin, glisin, triptofan ve izolösinden oluşan bir amino asit
karışımı, NH3:Benzen (10:90) hareketli fazı kullanılarak kağıt kromatografisi ile
ayrılmak isteniyor. Amino asitlerin yürüme hızlarının nasıl olmasını beklersiniz?
3- Amino asitler organizmada protein yapısına katılmaktan başka hangi görevleri
yaparlar?
Şekil 2.3- Protein yapısında yaygın olarak bulunan amino asitler.
Biyokimya Laboratuvarı I – Güz
10
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY III
AMİNO
ASİTLERİN
VE
PROTEİNLERİN
ÖZELLİKLERİNİN İNCELENMESİ
BAZI
I- AMİNO ASİTLER
Amino asitler yapılarında en az bir karboksil ve bir amino grubu içeren organik
bileşiklerdir. Tabiatta bulunan yaklaşık 300 amino asidin 20 tanesi protein yapısına girer.
Protein yapısına giren amino asitler L, α- aminoasitlerdir. Amino asitler peptid (amid)
bağları ile bağlanarak polipeptid ve proteinleri oluştururlar. Amino asitler polar
olmayan organik çözücülerde çok az, etanolde az, suda çok çözünürler ve nötral çözeltiler
verirler.
Hem asidik, hem de bazik çözeltilerde artan
bir çözünürlüğü, yani amfoter bir karaktere
sahiptirler. Erime ve bozulma noktaları 120300 °C aralığındadır.
Saf bir amino asidin suda çözülmesiyle
meydana gelen çözeltinin pH değerine
izoiyonik nokta denir. Amino asitlerin ve
proteinlerin
izoiyonik
noktada
çözünürlükleri maksimumdur.
8 amino asidi (valin, lösin, izolösin, treonin,
metiyonin, fenilalanin, triptofan, lizin)
organizma sentezleyemez. Bunlara esansiyel
amino asitler denir. Bu amino asitlerin
diyetle dışarıdan alınması gereklidir.
Şekil 3.1- Alanin amino asidinin stereoizomerleri
II- PEPTİDLER
Bir amino asidin amino grubu, ikinci bir amino asidin
karboksil grubu ile reaksiyona girdiğinde bir molekül
su ayrılması ile dipeptid oluşur. Arada oluşan kovalent
bağa peptid bağı denir ve rezonans gösterdiği için son
derece kararlıdır. 2 amino asit bir dipeptid, 3 amino asit
bir tripeptid, 10 veya daha az sayıda amino asit bir
oligopeptid, 50-100 amino asit ise bir polipeptid
oluşturur. Peptidlerin asidik ve bazik özellikleri
zincirin iki ucundaki amino ve karboksil grupları ile
yan zincirlerde bulunan iyonlaşabilen gruplara bağlıdır.
Biyokimya Laboratuvarı I – Güz
Şekil 3.2- Peptid bağının oluşumu
11
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 3.3- Serilglisiltirozilalanillösin pentapeptidi
Peptidler N-terminali (peptid zincirindeki serbest amino grubu) solda, C-terminali
(peptid zincirindeki serbest karboksil grubu) sağda kalacak şekilde yazılırlar ve bu
şekilde isimlendirilirler. Bu pentapeptid serilglisiltirozilalanillösin veya Ser–Gly–Tyr–
Ala–Leu olarak isimlendirilir.
III- PROTEİNLER
100’den fazla aminoasit içeren polipeptidlere protein adı verilir. Bazen belirli bir
biyolojik fonksiyonu gerçekleştirmek üzere, belirli bir konformasyonu almış olmak şartı
ile 100’den daha az sayıda amino asit içeren polipeptidlere de protein adı verilebilir.
Canlı hücrelerin kuru ağırlığının yarısından fazlasını proteinler oluşturur. Proteinler
organizmada enzimlerin, hormonların, reseptörlerin, bağışıklık sistemi ve pıhtılaşma
faktörlerinin yapısında bulunurlar. Yapı ve hareket sistemi içinde görev alırlar.
Amino asitler birleşerek peptid zincirini oluştururlar. Bu proteinlerin primer yapısıdır.
Peptid zincirleri daha kararlı bir hale gelmek üzere temel bir üç boyutlu yapı oluştururlar.
Buna proteinlerin sekonder yapısı denir. Bir polipeptidde birden fazla sekonder yapı bir
araya gelerek çeşitli kimyasal etkileşmeler sonucu üç boyutlu yapıda katlanırlar. Buna
tersiyer yapı denir. Birden fazla tersiyer yapıda katlanmış polipeptid zincirleri bir araya
gelerek kuaterner yapıyı oluştururlar.
Şekil 3.4- Proteinlerin yapısı
Proteinin doğal yapısındaki herhangi bir değişim denatürasyon olarak adlandırılır.
Sıcaklık, asitler (HCl, trikloroasetik asit, perklorik asit vb.) organik çözücüler (etanol,
aseton vb.), çapraz bağlayıcılar (formaldehit, glutaraldehit), kaotropik ajanlar (üre 6-8 M,
guanidin klorür 6M), indirgeyici ajanlar (2-merkaptoetanol, ditiyotreitol, iyodoasetat),
Biyokimya Laboratuvarı I – Güz
12
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
yüksek konsantrasyonda tuzlar (amonyum sülfat), ağır metal tuzları (cıva(II)klorür,
bakır(II)sülfat vb.) ve bazı deterjanlar (sodyumdodesilsülfat) proteinlerde denatürasyona
yol açarlar. Eğer denatüre edici faktörün ortamdan kaldırılması ile protein doğal yapısını
yeniden kazanabiliyorsa buna geri dönüşümlü denatürasyon denir. Eğer denatüre edici
faktörün ortamdan kaldırılması ile protein doğal yapısını yeniden kazanamıyorsa buna
geri dönüşümsüz denatürasyon denir.
Denatüre olan proteinlerde kuaterner, tersiyer ve kısmen sekonder yapılar bozulur ancak
primer yapı bozulmaz. Eğer polipeptid zincirinde kopmalar meydana geliyorsa buna
degradasyon adı verilir ve geri dönüşümsüzdür.
Düşük konsantrasyonlarda tuzlar proteinlerin çözünürlüğünü arttırırlar. MgCl2,
(NH4)2SO4 gibi iki değerlikli tuzlar, NaCl ve NH4Cl gibi tek değerlikli tuzlara göre daha
etkilidirler. Bu tuzların konsantrasyonları arttıkça proteinlerin çözünürlükleri azalır ve
yüksek tuz konsantrasyonlarında proteinler çöker.
KİMYASALLAR
Glisin, arjinin, glutamik asit, fenilalanin, tirozin, triptofan, histidin, sistein, sistin, kazein,
yumurta albümin, sülfat asidi, nitrat asidi, hidroklorik asit, sodyum hidroksit,
cıva(II)klorür, sodyumnitroprussiyat, bakır(II)sülfat, bakır(II)asetat, kadmiyum sülfat ve
bizmut(II)nitrat.
YÖNTEM
I- Amino Asitlerin Reaksiyonları
Amino asitlerin çözünürlüğü ve pH’sı:
4 ayrı tüpe az miktarda katı glisin, arjinin, glutamik asit ve fenilalanin alınır. Tüm
tüplere eşit miktarda destile su ilave edilir ve karıştırılır. Amino asitlerin çözünürlükleri
kaydedilir. Oluşan çözeltilerin pH’ları pH kâğıdı ile tayin edilir.
Ksantoprotein Reaksiyonu:
Fenil halkası içeren aromatik amino asitler derişik nitrat asidi ile muamele edildiklerinde
sarı renkli nitro türevlerine dönüşürler. Nitrat asidinin cilde temas etmesiyle oluşan
sararmanın sebebi budur.
Şekil 3.5- Ksantoprotein reaksiyonu
Biyokimya Laboratuvarı I – Güz
13
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Tirozin, triptofan, fenilalanin, histidin ve kazein’in sulu çözeltilerinden yaklaşık olarak az
miktarda alınarak üzerine 5 damla derişik nitrat asidi konur. Reaksiyona zor giren
fenilalanin tüpüne 5 damla daha nitrat asidi ve 5 damla derişik sülfat asidi eklenir. Tüpler
su banyosunda ısıtılır ve renk değişimi kaydedilir. Daha sonra tüpler bir portüpe alınarak
soğumaya bırakılır. Tüpler oda sıcaklığına geldiğinde %10 NaOH çözeltisi, ortam bazik
oluncaya kadar eklenir. Renk değişimi kaydedilir.
Millon Reaksiyonu:
Millon Reaktifi: 15 g HgCl2, 100 mL %50 HNO3 içinde çözülerek hazırlanır.
Fenol içeren bileşikler bu reaksiyonu verir. Tirozin amino asidi fenol grubu içeren tek
amino asit olduğundan bu reaksiyon tirozine özgüdür. Önce tirozinin fenol grubu reaktif
çözeltisindeki nitrik asit tarafından nitratlanır. Daha sonra nitratlanmış tirozin molekülü
çözeltideki cıva (I) ve cıva (II) iyonları ile kırmızı renk verir.
Ayrı tüplerdeki az miktarda tirozin ve kazeinin sulu çözeltilerine birkaç damla millon
reaktifi eklenir ve tüpler kaynar su banyosunda ısıtılır. Renk oluşumu kaydedilir.
Nitroprussiyat Reaksiyonu
Sülfidril grupları içeren bileşikler ağır metaller ve sodyum nitroprussiyat ile koyu renkli
kompleksler oluştururlar. Sistein serbest –SH grupları içerdiğinden bu reaksiyonu verir.
Sistin serbest –SH grubu içermediğinden bu reaksiyonu vermez.
Şekil 3.6- Sistin oluşumu
Şekil 3.7- Sodyum nitroprussiyat
3 tüpe ayrı ayrı sistein, sistin ve kazein çözeltileri alınır ve üzerlerine az miktarda %10
NaOH ve aynı hacimde %2 sodyum nitroprussiyat çözeltisi eklenir. Renk değişimi
kaydedilir. Kazein tüpüne bir miktar derişik HCl eklenir ve gözlemler kaydedilir.
Biyokimya Laboratuvarı I – Güz
14
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Triptofan Reaksiyonu
İki ayrı tüpteki az miktarda triptofan ve glisin çözeltileri üzerine eşit hacimde derişik
asetik asit konulur. Bu karışımın üzerine damla damla, tabaka oluşturacak şekilde derişik
sülfat asidi ilave edilir. Turuncu-mor halka oluşumu triptofan varlığını belirtir. Bu
triptofana özgü bir denemedir.
Aminoasitlerin verdikleri genel tanınma reaksiyonları:
Reaksiyon
Reaktif
Amino asit
Renk
Ninhidrin
Reaksiyonu
Ninhidrin
Serbest NH2 Grubu
Mor
Biüret
Alkali CuSO4
Amid Bağları
Mavi
Lowry Reaksiyonu
Fosfomolibdat
Tirozin
Mavi
Millon Reaksiyonu
HNO3, HNO2, HgNO3
Tirozin
Kırmızı
Ksantoprotein
Kaynar HNO3
Triptofan, Tirozin,
Fenilalanin
Sarı
Ehrlich Reaksiyonu
Der. HCl içinde pdimetilamino benzaldehit
Triptofan
Mavi
Sakaguchi
Reaksiyonu
α- Naftol, NaOCl veya
NaOBr
Arjinin
Kırmızı
Nitroprussiyat
Reaksiyonu
Sodyum nitroprussiyat,
NaOH
Sistein
Kırmızı
Sullivan reaksiyonu
Sodyum-1,2-naftokinon-4sülfonat ve NaHSO4
Sistein
Kırmızı
Kurşun Sülfür
Reaksiyonu
NaOH, Seyreltik Kurşun
asetat
Sistein
Siyah
Hopkins-Cole
Reaksiyonu
Glioksilik asit, Sülfürik Asit
Triptofan
Mor
Pauly Reaksiyonu
Sülfanilik asit, HCl, Sodyum
Histidin, Triptofan
karbonat, Sodyum nitrit
Biyokimya Laboratuvarı I – Güz
Histidin: Sarı
Triptofan: Mor
15
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
II- Proteinlerin Reaksiyonları
Proteinler ile ilgili denemelerde yumurta albümin çözeltisi kullanılacaktır. Yumurta
albümin çözeltisinin hazırlanması için bir yumurtanın akı, sarısından ayrılarak bir behere
aktarılır. Hacminin altı katı kadar su ilave edilir ve çözeltinin köpürmemesine dikkat
edilerek karıştırılır.
Proteinlerin Denatürasyonu:
Isının etkisi: Bir tüpe az miktarda yumurta albümin çözeltisi alınır ve bir süre sıcak su
banyosunda tutulur. Gözlemler kaydedilir.
Konsantre asitlerin etkisi: Bir tüpe az miktarda yumurta albümin çözeltisi alınır ve
üzerine hacminin iki katı kadar derişik nitrik asit ilave ederek gözlemler kaydedilir.
Ağır metal tuzlarının etkisi: 3 ayrı tüpe az miktarda yumurta albümin çözeltisi alınır ve
üzerlerine az miktarda ayrı ayrı bakır(II)asetat, kadmiyumsülfat ve bizmut(II)nitrat ilave
edilerek gözlemler kaydedilir.
Biüret reaksiyonu:
İki veya daha fazla peptid bağı ihtiva eden bileşikler alkali bakır sülfat ile menekşe veya
mor renkli kompleksler oluştururlar. Rengin koyuluğu proteindeki peptid bağlarının
sayısına bağlıdır. Bu test, peptid bağlarını belirlediğinden amino asitlerle reaksiyon
vermez. Bu reaksiyon ayrıca azot veya karbon atomuna bağlı iki karbonil grubu ihtiva
eden bileşiklerle de pozitif sonuç verir. Bu test proteinlerin kantitatif olarak tayini için de
kullanılmaktadır ancak bu amaçla kullanılan biüret reaktifi sodyum potasyum tartarat,
bakır sülfat ve potasyum iyodür içerir.
Üç ayrı tüpe az miktarda glisin, yumurta albümin ve kazein çözeltileri alınır. Tüplere aynı
hacimde %10 NaOH çözeltisi ve beşer damla %1 CuSO4 çözeltisi eklenir ve renk
değişimi kaydedilir.
Şekil 3.8- Biüret reaksiyonu sonucunda oluşan bakır-protein kompleksi
Biyokimya Laboratuvarı I – Güz
16
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Biyokimya Laboratuvarı I – Güz
17
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
ÇALIŞMA SORULARI
1- Glu-His-Trp-Ser-Gly-Leu-Arg-Pro-Gly dizisine sahip peptidi çiziniz. Bu peptidin
pH 3, 8 ve 11’deki net yükü nedir? (Bir grubun pKa değerinin altında ve üstünde
net yük değişiminin nasıl olduğunu hatırlayın). Bu peptidin pI değeri ne olabilir?
2- Organizmada görev yapan önemli peptidlere birkaç örnek veriniz ve ne görev
yaptıklarını belirtiniz.
Şekil 3.9- Amino asitlerin amino ve karboksil gruplarının pKa değerleri
Biyokimya Laboratuvarı I – Güz
18
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY IV
TAMPON ÇÖZELTİLER
Zayıf bir asidin kendisini ve anyonunu (konjuge bazını) veya zayıf bir bazın kendisini ve
katyonunu (konjuge asidini) yan yana içeren çözeltilere tampon çözeltiler adı verilir.
Tampon çözeltiler az miktarda asit veya baz ilavesiyle belirgin bir pH değişimi
göstermezler. Bir çözeltinin tampon etkisini gösterebilmesi için ayrı ayrı hem H+ iyonu
veren hem de H+ iyonu alan tanecikleri yeterli konsantrasyonda içermesi gereklidir.
Tampon çözeltiler sabit pH aralıklarında çalışmak üzere hazırlanırlar.
Henderson-Hasselbalch Denklemi
(A-: Tuz / HA: Asit)
Bu denklem bir çözeltinin teorik pH değerini hesaplamak için kullanılır. Denklemden de
görülebileceği gibi tamponu oluşturan maddeler arasında oran değişiklikleri yapılarak
pH’sı farklı tamponlar elde edilebilir.
Tampon çözeltiyi oluşturan maddeler her oranda değil sadece belirli bir oranda en iyi
tampon etkisini gösterirler. A-/HA oranı bire eşit olduğunda log1 = 0 olacağından pH
değeri de pKa değerine eşit olur. Bu pH değerine tamponun optimum pH değeri denir.
Bu pH değerinin dışındaki değerlerde tamponun etkisi zayıflar. Tamponlar optimum pH
değerlerinin bir birim altı ve bir birim üstündeki pH aralıklarında en etkili olarak
kullanılabilirler.
Tamponlar biyolojik ortamların hazırlanmasında faydalanılan çözeltilerdir. Canlı
organizmalarda pH değerleri son derece küçük bir aralıkta sabittir ve bu değerler sıkı bir
şekilde kontrol altında tutulur. Çünkü pH’daki değişiklikler enzimler, hücre membranları
ve nükleik asitler gibi birçok moleküler yapının yüklü bölgelerine etki eder ve fizyolojik
aktivitelerini etkiler. pH değerlerindeki 0.5 birimlik bir değişim canlının yaşamını
tehlikeye sokar. Organizmanın farklı bölgeleri farklı pH değerlerine sahiptir. (kan: 7.4,
mide özsuyu 1.5, pankreas özsuyu 8.0, beyin omurilik sıvısı 7.4 gibi). Bu bölgelerdeki
pH farklı tampon sistemlerince korunur. Bu sistemlere örnek olarak bikarbonat/karbonik
asit (HCO3-/H2CO3), protein/proteinat, oksihemoglobin/protonlanmış oksihemoglobin
(HbO2/HHbO2) ve fosfat (HPO4-/H2PO4-) tampon sistemleri verilebilir.
Organizma sıvılarının asit baz dengesinin asit tarafına kaymasına asidoz, alkali tarafına
kaymasına ise alkaloz denir. Asit metabolizma ürünleri artarsa, oluşan H+ iyonları HCO3/CO2 tarafından tutulur ve H2CO3 meydana gelir. Bunun dissosiasyonu ile oluşan H+
iyonları diğer tamponlarca tutulur. Bir kısmı da karbondioksit ve suya dönüşür.
Karbondioksidin artması nedeniyle pH 7.4’ün altına düşer. HCO3- konsantrasyonunun
azalması ile meydana gelen bu duruma metabolizma asidozu denir. Diyabette, açlıkta ve
karbonhidratsız rejimlerde görülen asidozlar buna örnektir. HCO3- konsantrasyonunun
artması ile metabolizma alkalozu meydana gelir. Aşırı sodyum bikarbonat alınması ve
şiddetli kusma sonucu midede HCl kaybı ile oluşan alkalozlar buna örnektir.
Biyokimya Laboratuvarı I – Güz
19
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
KİMYASALLAR
Monopotasyum fosfat (KH2PO4), disodyum fosfat (Na2HPO4), NaOH, HCl.
YÖNTEM
Tampon çözelti hazırlama tablolarından yararlanarak 50 mL pH 7.4 Fosfat (Sørensen),
25 mL pH 6 sitrikasit/fosfat tamponlarını hazırlayınız. Hazırladığınız tamponun pH
değerini ölçünüz. Eğer pH değerleri gerekenden farklı bir değere sahip ise 1N NaOH
veya 1N HCl kullanarak pH’sını ayarlayınız.
ÇALIŞMA SORULARI
1- Bir tampon litrede 0.01 mol laktik asit (pKa=3.86) ve 0.05 mol sodyum laktat
içermektedir.
a- Tamponun pH değerini hesaplayınız.
b- 1 L tampona 5 mL 0.5 M HCl eklendiğinde pH değişimi ne olur?
2- Organizma sıvılarında pH düzenlemesinde en önemli görev üstlenen organlar
nelerdir ve bunu hangi yollarla gerçekleştirirler.
Biyokimya Laboratuvarı I – Güz
20
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Biyokimya Laboratuvarı I – Güz
21
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Biyokimya Laboratuvarı I – Güz
22
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY V
KATEKOLÜN OKSİDASYONUNA
OKSİDAZ ENZİMİNİN ETKİSİ
POLİFENOL
Enzimler canlı organizmalardaki reaksiyonları katalizlerler. Laboratuvar ortamında çok
yüksek veya çok düşük basınç, pH ve sıcaklıkta oluşan kimyasal değişme ve
reaksiyonlar, canlı hücrelerin bozulmadan kalabildikleri dar basınç, pH ve sıcaklık
aralığında, ancak enzimler aracılığıyla gerçekleştirilebilirler.
Küçük bir grup katalitik RNA moleküllerinin haricinde tüm enzimler protein
yapısındadırlar. Katalitik aktiviteleri genellikle proteinin doğal yapısında olmasına
bağlıdır. Eğer bir enzim denatüre olur veya alt ünitelerine ayrılırsa katalitik aktivitesi
genellikle kaybolur.
Bazı enzimler aktivite göstermek için bir veya daha fazla anorganik iyona veya kompleks
organik veya metallorganik bileşiklere ihtiyaç gösterirler. Bu iyon ya da bileşiğe
kofaktör adı verilir. Organik bileşik enzimin protein kısmı ile çok sıkı bir şekilde
bağlanmış ise prostetik grup, çok sıkı bağlanmamış ve dissosiye olabiliyorsa koenzim
adını alır. Katalitik olarak etki gösteren tam enzime holoenzim denir. Holoenzimin
protein kısmına ise apoenzim adı verilir.
Şekil 5.1- Enzim yapısı
Enzimin etki ettiği bileşiğe substrat adı verilir. Enzimin katalizlediği reaksiyonu
proteinin tamamı değil, sadece “aktif bölge” adı verilen belirli bir bölgesi etkiler.
E=Enzim, S= Substrat, ES= Enzim-Substrat kompleksi ve P= Ürün olmak üzere enzim
reaksiyonu şu şekilde özetlenebilir.
Biyokimya Laboratuvarı I – Güz
23
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Böyle bir reaksiyon genel kimyasal reaksiyon mekaniğine ve kinetiğine uyar. Diğer
katalizörlerde de olduğu gibi enzimler reaksiyonun yönünü değiştirmezler sadece
reaksiyonun hızına etki ederler. Reaksiyona ilerleyebileceği daha düşük aktivasyon
enerjili ara yollar sunarak reaksiyonu hızlandırırlar.
Şekil 5.2- Enzimli reaksiyonun serbest enerji diyagramı
Şekilde ΔGEnzimsiz enzimsiz reaksiyonun aktivasyon enerjisi,
reaksiyonun aktivasyon enerjisidir. ΔH Reaksiyon entalpisidir.
ΔGEnzimli ise enzimli
Reaksiyon başında substrat konsantrasyonu yüksek, ürün konsantrasyonu ise ihmal
edilecek kadar azdır. ES kompleksi büyük bir serbest enerji düşmesi ile ayrıldığından k4
ihmal edilebilir. Öyleyse ES oluşumunun net hızı, ES bozunmasının net hızına eşit
olacaktır ki denklem 1’e göre bu eşitlik;
olur.
Reaksiyonun başında [P] ihmal edilebilir olduğundan,
şeklinde yazılabilir.
Burada Km, kontrollü şartlarda reaksiyonun maksimum hızının yarısına eşit hız veren
substrat konsantrasyonudur ve Michaelis-Menten sabiti olarak adlandırılır.
Biyokimya Laboratuvarı I – Güz
24
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
[E0], başlangıçtaki toplam enzim konsantrasyonu ve reaksiyonun herhangi bir anındaki
enzim konsantrasyonu [E] ise, [E] = [E0] – [ES] olur.
Denklem 3 aşağıdaki şekle dönüşür;
Maksimum hız toplam enzim konsantrasyonu ile orantılı olduğu kadar, başlangıç hızıyla
da orantılıdır. Böylece Vmaks = K3 [E0] olduğundan denklem 4’te yerine konulursa
aşağıdaki denklem elde edilir.
Denklem 5’te başlangıç hızı V0 yalnız bırakılacak olursa;
eşitliği bulunur.
Denklem 6’ya göre V0 ile [S] arasında çizilen grafik aşağıdaki gibi olur. Bu eğriye
Michaelis-Menten eğrisi denir.
Şekil 5.3- Michaelis-Menten eğrisi
Biyokimya Laboratuvarı I – Güz
25
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Düşük substrat konsantrasyonlarında denklem 6’nın paydasında Km >> [S] olduğunda
[S], Km yanında ihmal edilebilir ve = Vmaks [S] / Km olur. Yüksek substrat
konsantrasyonlarında denklem 6’nın paydasında Km << [S] olacağından Km, [S] yanında
ihmal edilebilir, V0= Vmaks olur ve denklem 6’daki eşitlikle bölünerek doğru denklemi
verecek şekle dönüştürülebilir.
Y eksenine 1/V0, X eksenine de 1/[S] değerleri yerleştirilerek çizilen grafikte elde edilen
doğrunun eğimi Km/Vmaks’a ve Y eksenini kesen nokta 1/Vmaks’a eşit olur. Km değeri de
grafikte gösterildiği şekilde bulunabilir. Bu eğriye Lineweaver-Burk eğrisi adı verilir.
Eğer V0 değerine karşı V0/[S] grafiği çizilecek olursa bu eğriye Eadie-Hofstee eğrisi
elde edilir.
Km değerlerinden enzimlerin ayırt edilmesinde, ES komplekslerine ilişkin sabitlerin
bulunmasında faydalanılır Km substrat konsantrasyonu birimi cinsindendir (mol/L veya
mmol/L).
Şekil 5.4- Lineweaver-Burk eğrisi
Şekil 5.5- Eadie-Hofstee eğrisi
Enzim aktivitesinin ölçülmesinde çeşitli birimler kullanılır. En çok kullanılan aktivite
birimi enzim ünitesidir (aktivite).
Enzim ünitesi (U): 25°C’de 1 dakikada optimum şartlarda 1µmol substratı ürüne çeviren
enzim miktarıdır.
Spesifik aktivite (U/mg protein): 1 mg protein başına enzim ünitesidir. Enzimin saflık
derecesinin bir göstergesidir.
Molar Aktivite: Bir tek enzim molekülü tarafından birim zamanda ürüne çevrilen
substrat molekülü sayısıdır. Dönüşüm sayısı olarak da adlandırılır.
Katal: 1 saniyede 1 mol substratı reaksiyona sokan enzim miktarıdır.
Biyokimya Laboratuvarı I – Güz
26
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Enzim miktarı çok küçük olduğundan direkt olarak ölçmek zordur. Optimum şartlarda
katalizledikleri reaksiyon hızları diğer şartlar sabit kalmak üzere, enzim konsantrasyonu
ile orantılıdır. Böylece enzim konsantrasyonu hız ölçülmesi ile tayin edilebilir.
Enzimle katalizlenmiş reaksiyonların hızı inhibitör adı verilen bazı maddeler tarafından
azaltılır veya tamamen durdurulur. Buna enzim inhibisyonu denir. Enzim inhibisyonu
geri dönüşümlü veya dönüşümsüz olabilir.
Polifenol oksidaz enzimi doğada yaygın olarak bulunan, bakır içeren bir enzimdir. Meyve
ve sebzelerde hasar görmüş bölgelerin hava ile teması sonucu kararması ve cildin
esmerleşmesi (melanin sentezi) gibi reaksiyonlardan sorumludur. Enzimin aktivitesi,
fenollerin katekollere orto-hidroksilasyonu ve katekollerin orto-kinonlara oksidasyonu
olacak şekilde ikiye ayrılabilir.
Aktif merkezde substratın bağlanabileceği bir boşluk vardır ve bu merkezdeki histidin
kalıntıları bakır bağlamada görev yaparlar.
Şekil 5.6- Polifenol oksidaz enziminin 3 boyutlu yapısı ve aktif merkezi
Bu denemede katekolün benzokinona enzimatik dönüşüm reaksiyonunun kinetikleri
incelenecektir.
Biyokimya Laboratuvarı I – Güz
27
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 5.7- Polifenol oksidaz enziminin katalizlediği oksidasyon reaksiyonunun mekanizması
Bu reaksiyonda kullanılan katekol çözeltisi renksizdir, oluşan 1,2-benzokinon ise
kahverengidir. Böylece oluşan ürünün miktarı kolorimetrik olarak izlenebilir. 1,2benzokinon 480 nm dalga boyunda maksimum absorbans verdiğinden bu dalga boyunda
yapılan spektrofotometrik ölçüm yardımı ile belli bir zamanda çözeltide bulunan ürün
miktarı hesaplanabilir.
Şekil 5.8- Spektrofotometrenin çalışma prensibi
Spektrofotometrik ölçümde bir ışık kaynağı geniş bir spektrumda ışık yayar,
monokromatör belirli bir dalga boyundaki ışığı seçer ve örnek küvetine gönderir. Işık
küvetten geçerken absorblayıcı moleküllerin konsantrasyonuna bağlı olarak bir kısmı
absorblanır ve çözeltiden geçen ışık bir dedektör tarafından ölçülür.
Biyokimya Laboratuvarı I – Güz
28
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Bu denemede amaç iki anahtar parametrenin bulunmasıdır. Bunlar maksimum hız (Vmaks)
ve Michaelis-Menten sabitidir (Km).
KİMYASALLAR
Sitrik asit-fosfat tamponu pH 6, katekol, benzokinon.
YÖNTEM
Ham enzimin hazırlanması: Patatesler soyulur ve bıçakla küçük parçalara ayrılır. Daha
sonra blender cihazı yardımıyla ve 200 mL pH 6 sitrik asit - fosfat tamponu varlığında
homojenize edilir. Enzimlerin aktivitesi sıcaklıktan etkilendiği için bu aşamadan
sonraki tüm işlemler hızlı bir şekilde ve buz üzerinde gerçekleştirilir. Elde edilen
homojenizat, bir buz banyosuna önceden yerleştirilerek soğutulmuş erlen ve huni
yardımıyla cam pamuğundan süzülür ve süzüntü deney süresince buz banyosunda tutulur.
Substrat konsantrasyonunun reaksiyon hızına etkisi: 6 adet deney tüpü aşağıdaki
tabloda gösterildiği şekilde (enzim konulmadan) hazırlanır. Enzim çözeltisi
spektrofotometrik ölçümden hemen önce konulacaktır.
Tüp
No
Substrat
Katekol
0.01 M
(mL)
Su
(mL)
Enzim
(µL)
1
2
3
4
5
6
7
0
1
2
3.5
5
7
9
10
9
8
6.5
5
3
1
500
500
500
500
500
500
Absorbans
1. dk.
2. dk
3. dk
4. dk
5.dk
6.dk
Grafikten
Hesaplanan
Hız
Sıfırlama yapılır
2-7 numaralı tüplerin dakika başına verdikleri absorbans değeri ölçülecektir. Bu işlem
şöyle yapılır;
1 numaralı tüpe 500µL enzim konulur ve tüp iyice karıştırılarak içeriği iki
spektrofotometre küvetine aktarılır. Bu çözeltiler bizim şahit çözeltilerimiz olacaktır. İki
şahit küveti kullanılarak spektrofotometre 480 nm dalga boyunda sıfırlanır. Daha sonra 2
numaralı tüpe enzim konulur ve enzim konulduğu anda kronometre çalıştırılır. İçinde
enzim bulunan 2 numaralı tüp iyice karıştırılır, içeriği bir spektrofotometre küvetine
aktarılır ve spektrofotometreye yerleştirilir.
Biyokimya Laboratuvarı I – Güz
29
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Bu sırada kronometre takip edilerek ilk enzimin konulmasından tam 30 saniye sonra 3
numaralı tüpe 500 µL enzim konulur ve iyice karıştırılarak içeriği bir spektrofotometre
küvetine aktarılır ancak spektrofotometreye yerleştirilmez. Kronometre 01:00 değerini
gösterdiğinde
spektrofotometrenin
gösterdiği
absorbans
değeri
kaydedilir.
Spektrofotometrenin içindeki 2 numaralı çözeltiyi içeren küvet çıkarılır ve 3 numaralı
çözeltiyi içeren küvet yerleştirilir. Kronometredeki değer 01:30 değerini gösterdiğinde
spektrofotometrenin gösterdiği absorbans değeri kaydedilir. Küvetler değiştirilerek her
bir küvet için dakikada bir absorbans değeri ölçülür. Her bir küvet için 6 değer alınıncaya
kadar bu işlem devam eder. Daha sonra aynı işlemler 4-5 ve 6-7 numaralı tüpler için de
tekrarlanır. Her işlemin başında spektrofotometre şahit çözeltiler kullanılarak sıfırlanır.
Ekstinksiyon sabitinin bulunması: 100 mL 0.01 M benzokinon çözeltisi hazırlanarak
480 nm dalga boyundaki absorbansı ölçülür. Benzokinon zor çözündüğünden bu
çözelti deney başladığında hazırlanır ve ölçümü deneyin sonunda yapılır.
VERİLERİN KULLANIMI
2-7 numaralı tüpler için 480 nm’de elde edilen absorbans değerleri ayrı ayrı, zamana
karşı grafiğe çizilir.
Şekil 5.9- Absorbans- zaman grafiği
Her grafikte, hiperbolik eğrilere teğet doğrular çizilir ve bu doğruların eğimleri
hesaplanır. Eğim = ΔA/Δt, A=ε.c.l olduğundan her bir küvet için başlangıç hızı
V = m / ε . l denkleminden hesaplanır.
2-7 numaralı tüplerin her biri için katekol konsantrasyonları hesaplanır. Bulunan
başlangıç hızları ile substrat konsantrasyonları arasında Michaelis-Menten eğrisi çizilir.
Bu hız ve substrat konsantrasyonları için Lineweaver-Burk ve Eadie-Hofstee grafikleri
çizilerek Vmaks ve Km değerleri hesaplanır.
Biyokimya Laboratuvarı I – Güz
30
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
ÇALIŞMA SORULARI
1- Substrat ile doygun olduğu zaman 10-6 M konsantrasyonundaki karbonik anhidraz enzimi
saniyede 0.6 mol/L karbonik asit oluşumunu katalizler. Buna göre dönüşüm sayısı nedir.
2- Geri dönüşümlü enzim inhibisyonu, inhibitörün enzime bağlanmasına göre yarışmalı,
yarışmasız ve yarı-yarışmalı olmak üzere üçe ayrılabilir. Aşağıda verilen şekilleri ve
grafikleri inceleyerek her bir durum için inhibitör konsantrasyonu değişimi ile V maks ve
Km değerlerinin nasıl değiştiklerini yorumlayınız.
Şekil 5.10- Enzim inhibisyon mekanizmaları
Biyokimya Laboratuvarı I – Güz
31
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY VI
PROTEİNLERİN SAFLAŞTIRILMASI VE KARAKTERİZASYONU
I- Proteinlerin Ekstraksiyonu ve Diyaliz
Bir proteinin yapısının ve özelliklerinin araştırılabilmesi için, o proteinin saf bir şekilde
elde edilmesi gerekir. Proteinler kolaylıkla denatüre olabildiklerinden ve biyolojik
materyallerde (örneğin vücut sıvılarında ve doku ekstraktlarında) kompleks bir karışım
halinde bulunduklarından, saflaştırılmaları oldukça zordur.
Proteinlerin elde edilebildiği kaynaklar genellikle dokular ve hücrelerdir. Bir proteinin
saflaştırılmasında yapılacak ilk işlem hücrelerin parçalanarak içerdikleri proteini bir
çözeltiye almaktır. Bu çözeltiye ham ekstrakt denir. Bu ham ekstrakt istenen proteinin
yanı sıra pek çok başka protein ve molekül de içermektedir.
Daha sonra proteinlerin çözünürlük farkları, büyüklükleri, elektriksel yükleri ve
polariteleri gibi çeşitli özelliklerinden yararlanılarak protein ekstraktı fraksiyonlara
ayrılır. Bu özelliklere dayanarak proteinlerin saflaştırılması ile ilgili pek çok yöntemler
kullanılmaktadır. Bunlara örnek olarak kademeli santrifüj, tuzla çöktürme, aseton ve
alkolde soğukta çöktürme, ultrafiltrasyon ve çeşitli kromatografik yöntemler verilebilir.
Aşağıda ham ekstrakttan bir enzimin kromatografik yöntemler kullanılarak saflaştırılması
için örnek bir yöntem verilmiştir.
abcde-
Ham ekstraktın elde edilmesi
Tuzla (amonyum sülfat) çöktürülmesi
İyon değişim kromatografisi
Jel geçirgenlik (moleküler elek) kromatografisi
Afinite (ilgi) kromatografisi.
Proteinler saflaştırma işlemi sırasında her bir basamakta çeşitli kayıplara uğrarlar ve
aktiviteleri azalır. Ancak istenilen enzim her bir basamakta daha saf hale gelir, bu
nedenle daha derişik hale gelir ve spesifik aktivitesi artar. Enzimlerle çalışıldığında
ortamın pH’sı ve sıcaklığının çok iyi bir şekilde kontrol edilmesi gerekir.
İşlem Basamağı
Fraksiyon Hacmi
(mL)
Total Protein
(mg)
Aktivite
(Ünite)
Spesifik Aktivite
(Ünite/mg)
Ham Ekstrakt
1400
10000
100000
10
Tuzla Çöktürme
280
3000
96000
32
İyon Değişim
90
400
80000
200
Jel Geçirgenlik
80
100
60000
600
Afinite
6
3
45000
15000
Biyokimya Laboratuvarı I – Güz
32
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
İyon Değişim Kromatografisi
Şekil 6.1- İyon değişim kromatografisi
Biyokimya Laboratuvarı I – Güz
33
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Jel Geçirgenlik (moleküler Elek) Kromatografisi
Şekil 6.2- Jel geçirgenlik (moleküler elek) kromatografisi
Biyokimya Laboratuvarı I – Güz
34
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Afinite (İlgi) Kromatografisi
Şekil 6.3- Afinite (ilgi) kromatografisi
Biyokimya Laboratuvarı I – Güz
35
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Tuzla çöktürme işlemi sonunda ortamda bulunan tuzlar saflaştırma işleminin ileri
safhalarında ve protein karakterizasyonu işlemleri sırasında deneyleri bozarlar. Bu
nedenle tuzların ortamdan uzaklaştırılması gerekir. Bu amaçla jel filtrasyon, ultra
santrifüj ve diyaliz yöntemleri kullanılabilir. Büyüklük farkına dayanan bir ayırma
yöntemi olan diyaliz basit ve pahalı olmayan bir işlemdir.
Şekil 6.4- Diyaliz
Diyaliz işleminde yarı geçirgen selüloz membranlar kullanılır. Bu membranların gözenek
büyüklükleri bellidir ve sadece belli bir boyuttaki moleküllerin geçmesine izin verirler.
Diyaliz tamponu olarak tuz konsantrasyonu düşük olan çözeltiler kullanılır. Membranın
içindeki tuz konsantrasyonu, membranın dışındaki diyaliz tamponunun tuz
konsantrasyonundan daha fazla olduğu için membran içindeki tuzlar membran dışına
doğru hareket ederler. Proteinlerin molekül ağırlıkları tuzlar ile kıyaslandığında çok daha
büyük olduğundan membranın gözeneklerinden geçemezler ve membran içinde kalırlar.
Bu işlem diyaliz tamponu ve membran içindeki çözelti arasında bir tuz konsantrasyonu
dengesi oluşuncaya kadar devam eder. Bu dengenin oluşması yaklaşık 4–6 saat kadar
sürer. Dengeye ulaşıldığında diyaliz tamponu tazelenir, böylece denge bozulur ve tuzlar
membran dışına çıkmaya devam eder. Bu şekilde protein molekülleri tuzdan kurtarılmış
olur.
Biyokimya Laboratuvarı I – Güz
36
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
KİMYASALLAR
Potasyumdihidrojen fosfat, disodyumhidrojen fosfat, amonyum sülfat, üre, tiyoüre,
trishidroksimetil aminometan, diyaliz membranı.
YÖNTEM
Bu deneyde 3 farklı dokudan protein ekstraksiyonu yapılacaktır. Bu amaçla dana
karaciğeri, yumurta akı ve yumurta sarısı protein kaynakları olarak kullanılacaktır.
Liziz tamponu: 9 M üre, 0.04 M Tris ve %0.1 SDS çözeltisinden oluşur. 0.484 g Tris ve
54.63 g üre 100 mL’lik bir balonjojeye alınır ve pH 7.4 Sørensen tamponu ile hacim
tamamlanır. Daha sonra homojenizata 1 mL %10 SDS çözeltisi ilave edilir.
Homojenizasyon
Karaciğer yıkanır, parçalara ayrılır ve liziz tamponu varlığında blender cihazı
kullanılarak daha küçük parçalar haline getirilir. Daha sonra homojenizatör yardımı ile
homojenize edilir. Homojenizata 1 mL %10 SDS çözeltisi ilave edilerek karıştırılır. ve
santrifüj tüplerine alınarak santrifüj edilir. Süpernatant (tüpün üst kısmındaki sulu çözelti)
dekante edilerek bir behere alınır.
Yumurta akı ve sarısı sıvı oldukları için bu homojenizasyon basamağına gerek
duymazlar. Behere alınan yumurta akı ve sarısı üzerine liziz tamponu ve az miktarda %10
SDS ilave edilir. Köpürmemesine dikkat edilerek karıştırılır ve santrifüj edilerek
süpernatantlar ayrı ayrı bir behere toplanır.
Çöktürme
Tuzla çöktürme: Homojenize edilmiş olan çözeltilerden 10 mL alınır ve aşağıda verilmiş
olan doygunluk hesabı tablosundan yararlanılarak istenen amonyum sülfat
doygunluğunda çöktürme yapmak için gerekli olan amonyum sülfat miktarları hesaplanır.
Biyokimya Laboratuvarı I – Güz
37
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Gerekli miktar tartıldıktan sonra, katı amonyum sülfat protein örneklerini içeren tüplere
eklenir, dikkatle karıştırılır ve santrifüjlenir. Santrifüj sonunda süpernatant dekante
edilerek atılır ve presipitat (çökmüş olan katı kısım) üzerinden deneye devam edilir.
Asetonla çöktürme: Homojenize edilmiş olan çözeltilerden 4 mL alınır ve üzerine 8 mL
soğuk aseton ilave edilir. Dikkatle karıştırılır ve santrifüj edilir. Santrifüj sonunda
süpernatant dekante edilerek atılır ve presipitat üzerinden deneye devam edilir.
Çözünürleştirme
Çöktürülmüş olan proteinlere az miktarda pH 7.4 Sørensen fosfat tamponu ilave edilir.
Dikkatle karıştırılarak proteinlerin mümkün olduğunca çok çözünmesi sağlanır. Daha
sonra tüpler santrifüj edilerek süpernatantlar alınır ve presipitatlar atılır.
Diyaliz
Tuzla çöktürülmüş olan proteinler daha önceden hazırlanmış olan diyaliz torbalarına,
torbanın en fazla 3/4’ü dolacak şekilde yerleştirilir ve diyaliz tamponu olarak destile su
kullanılarak diyaliz edilirler. 4 saate bir diyaliz tamponu değiştirilerek soğukta 2 gün
boyunca diyaliz edilirler. Diyaliz sona erdikten sonra diyaliz torbaları açılır ve
diyalizatlar temiz bir tüpe alınır.
Bu işlemler sonrasında elde edilen çözeltiler protein örnekleridir, proteinlerin
konsantrasyon tayini ve elektroforez denemelerinde bu çözeltiler kullanılacaktır. Bu
amaçla elde edilen protein çözeltileri hacimlere ayrılır ve -20°C soğutucuda saklanırlar.
ÇALIŞMA SORULARI
1- Günlük hayatta saflaştırılmış proteinler nerelerde kullanılmaktadır? Araştırınız.
Biyokimya Laboratuvarı I – Güz
38
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY VII
PROTEİNLERİN KANTİTATİF TAYİNİ
Protein saflaştırılması işleminde her basamakta elimizde bulunan protein
konsantrasyonunun bilinmesi çok önemlidir. Sonraki basamaklarda hangi yöntemlerin
kullanılacağı protein konsantrasyonuna bağlı olarak belirlenir.
Çözünür proteinlerin çözelti içindeki konsantrasyonlarının ölçümü için birçok farklı
yöntem geliştirilmiştir. Her yöntemin bazı avantaj ve dezavantajları vardır. En yaygın
kullanılan 3 yöntem; basit absorbans yöntemi, Lowry yöntemi ve Bradford
yöntemidir. Bu yöntemlerde bilinen konsantrasyonlardaki protein çözeltileri ile
hazırlanan standart kalibrasyon grafikleri kullanılarak bilinmeyen örneğin
konsantrasyonu bulunur.
Her üç yöntem de proteinlerin analizi için spektrofotometrik ölçümleri kullanır.
Basit Absorbans Yöntemi
Bu yöntem proteinlerdeki tirozin ve triptofan amino asitlerinin 280 nm’de maksimum
absorbans vermesine dayanır. Bu absorbsiyona fenilalanin amino asidi ve disülfit bağları
da az miktarda olmakla birlikte katkıda bulunur.
Çok hızlı ve ucuz bir yöntemdir, az miktarda materyale ihtiyaç gösterir ayrıca protein
örneği üzerinde herhangi bir kimyasal modifikasyon yapılmadığından örnek geri
kazanılabilir. Ancak hassasiyeti düşüktür (0.05–2 mg protein/mL). Ayrıca farklı
miktarlarda tirozin ve triptofan içeriğine sahip proteinler aynı konsantrasyonda olsalar
bile farklı absorbans verirler. Yakın bir dalga boyunda maksimum absorbans
verdiklerinden nükleik asitler ve başka kromofor gruplarda etkileşim yapabilir.
Ayrıca uzak UV alanda da (190 nm) bu yöntem uygulanabilir. Peptid bağlarına bağlı
olarak absorbans verildiğinden amino asit kompozisyonunun değişiminden bağımsızdır
ve hassasiyeti yüksektir (0.01–0.05 mg). Ancak pek çok çözelti ve tampon bu bölgede
absorbans verdiğinden girişimler de artar. Ayrıca oksijen absorbsiyonundan ötürü
özelleşmiş cihazlara ihtiyaç duyar.
Sığır serum albümin (BSA) çözeltisi bu denemede sıklıkla standart olarak kullanılır.
Lowry (Folin-Ciocalteau) Yöntemi
Bu yöntem hem biüret hem de folin reaksiyonuna dayanır. Biüret reaksiyonunda alkali
şartlarda bakır, proteinlerin peptid bağları ile reaksiyona girerek Cu+ oluşturur. FolinCiocalteau reaksiyonunda ise fosfomolibdotungstat, aromatik amino asitlerin Cu katalizli
oksidasyonu ile heteropolimolibden mavisine indirgenir. Sonuçta kısmen tirozin ve
triptofan bileşimine bağlı olan, şiddetli koyu bir mavi renk elde edilir.
Biyokimya Laboratuvarı I – Güz
39
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Lowry yöntemi de tirozin ve triptofan içeriğine bağlıdır ancak proteinden proteine olan
farklılıklar orta seviyededir. 0.01 mg/mL hassasiyete sahiptir ve 0.01–1 mg/mL protein
içeren çözeltiler için kullanımı uygundur.
Bu yöntemin bir dezavantajı girişimlerin fazla oluşudur. Çeşitli tamponlar, ilaç aktif
maddeler, deterjanlar, EDTA, amonyum sülfat, Glisin, merkaptoetanol, nükleik asitler ve
şekerler gibi bir grup madde deneme ile girişim gösterir. Hücre ekstraktları ve kompleks
örnekler, örneğin perkloroasetik asit/ etanol çöktürmesi gibi bir temizleme basamağına
ihtiyaç gösterebilir. Günümüzde denemenin ihtiyaçlarına göre modifiye Lowry
yöntemleri kullanılmaktadır. Reaksiyon pH bağımlıdır, pH’nın 10–10.5 arasında olması
önemlidir. İnkübasyon basamağındaki süre kritik değildir 10 dakika ile birkaç saat
arasında değişebilir. Tüplerin iyi karıştırılması çok önemlidir, çünkü folin reaktifi alkali
ortamda kararsız olduğu için kısa bir süre aktivite gösterir. Deneme yüksek
konsantrasyonlarda lineer değildir, kalibrasyon eğrisinin buna göre hazırlanması gerekir.
Bradford Yöntemi
Bradford yöntemi Coomassie Brilliant Blue G250 (CBB) boyar maddesinin proteinlere
kovalent olmayan bağlanmasına dayanır. CBB’nin asidik çözeltideki proteine bağlanması
boyanın 465 nm’deki maksimum absorbsiyonunu 595 nm’ye kaydırır. 595 nm’deki
absorbsiyon doğrudan protein konsantrasyonlarına aittir.
Bu yöntem Lowry yöntemine göre daha hızlı, daha ucuz, daha kolay ve daha hassastır.
CBB’nin protein örneğine katılması ile 2 dakika gibi kısa bir zamanda renk oluşu ve bu
renk 1 saat sabit olarak kalır. 0.001 mg/mL protein ölçülebilir.
Lowry ile karşılaştırıldığında, yaygın kimyasal bileşikler ve biyolojik örneklerdeki
protein olmayan moleküllerden daha az etkilenir. Amfolitler, deterjanlar bu yöntemde
girişim yaparlar. CBB arjinin ve lizin amino asitlerine daha kolay bağlanır bu sebeple
farklı proteinler farklı renk yoğunluğu verebilirler. Bu yöntemin dezavantajlarından
biridir. CBB serbest arjinin ve lizine, 3000 kDa’dan daha küçük peptid moleküllerine
bağlanmaz.
Biyokimya Laboratuvarı I – Güz
40
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Yukarıdaki tablodan da görüldüğü gibi her kantitatif protein tayin yönteminin kendine
özgü avantajları ve dezavantajları bulunmaktadır. Bu sebeple bir protein tayini yöntemi
seçilirken yapılan çalışmanın özellikleri göz önünde bulundurulmalı ve çalışma
ortamındaki moleküllerle girişim yapmayacak, yeterli hassasiyette, ucuz ve uygulanabilir
bir yöntem seçilmelidir.
KİMYASALLAR
NaCl, Sığır Serum Albümin (BSA), Na2CO3, CuSO4·5H2O, KNaC4H4O6·4H2O, Folin
Reaktifi, Bradford Reaktifi, NaOH, Ovalbümin.
Biyokimya Laboratuvarı I – Güz
41
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
YÖNTEM
Basit absorbans
Serum Fizyolojik: %0.0897’lik NaCl çözeltisi hazırlanır.
BSA Stok Çözeltisi: Serum fizyolojik içinde %0.2’lik 100 ml BSA çözeltisi hazırlanır.
Aşağıdaki tabloya göre tüpler hazırlanır. Deneydeki hata oranını azaltmak ve
tekrarlanabilirliği arttırmak için tüpler çift çalışılmalıdır. Serum fizyolojik çözeltisi şahit
olarak kullanılarak her bir tüpün 280 nm’deki absorbansı ölçülür, kaydedilir ve protein
konsantrasyonları 2–5 numaralı tüpler için hesaplanır. Bu konsantrasyon verileri
kullanılarak kalibrasyon grafiği çizilir. Bilinmeyen örneğin absorbansı grafik üzerine
yerleştirilerek konsantrasyonu bulunur.
Tüp no
BSA Stok
(mL)
Serum
Fizyolojik
(mL)
1
0
3.0
2
0.5
2.5
3
1.0
2.0
4
1.5
1.5
5
2.0
1.0
6
Absorbans
(280 nm)
Protein
Konsantrasyonu
(µg/mL)
Bilinmeyen örnek (3.0)
Biyokimya Laboratuvarı I – Güz
42
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Lowry
Kompleks oluşturucu reaktif: Stok çözeltilerden taze olarak hazırlanmalıdır. 50 mL
%2’lik Na2CO3 (Çözelti A), 10 mL %1’lik CuSO4·5H2O (Çözelti B) ve 10 mL %2’lik
KNaC4H4O6·4H2O (Çözelti C) stok çözeltileri hazırlanır. Bu çözeltiler A:B:C 100:1:1
oranında karıştırılarak hazırlanır.
Protein stok çözeltisi: Serum fizyolojik içerisinde 2 mg/mL BSA çözeltisi hazırlanır. Bu
çözelti protein stok çözeltisi olarak kullanılacaktır.
Folin reaktifi: 1N Fosfomolibdat ve fosfotungstik asit çözeltisi (Bu çözelti hazır halde
verilecektir.)
Serum
Fizyolojik
(µL)
%8’lik
NaOH
Çöz.(µL)
1
0
500
500
2
25
475
500
3
50
450
500
4
75
425
500
5
100
400
500
6
Bilinmeyen Örnek
(500)
500
Komp.
Oluş.
Reakt.
(µL)
1000
1000
1000
1000
1000
1000
Biyokimya Laboratuvarı I – Güz
Folin
reaktifi
100
100
100
100
100
100
İyice karıştırılır ve 30 dk. İnkübe edilir
Protein
Stok
Çöz.
(µL)
Karıştırılır ve 10 dk. inkübe edilir
Tüp
No
100°C’de 10 dk. inkübe edilir ve oda
sıcaklığına soğutulur
Aşağıdaki tabloya göre tüpler hazırlanır. Deneydeki hata oranını azaltmak ve
tekrarlanabilirliği arttırmak için tüpler çift çalışılmalıdır. 1 numaralı tüp şahit olarak
kullanılarak her bir tüpün 550 nm’deki absorbansı ölçülür, kaydedilir ve protein
konsantrasyonları 2-5 numaralı tüpler için hesaplanır. Bu konsantrasyon verileri
kullanılarak kalibrasyon grafiği çizilir. Bilinmeyen örneğin absorbansı grafik üzerine
yerleştirilerek konsantrasyonu bulunur.
Absorb.
(550
nm)
Protein
Kons.
(µg/mL)
43
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Bradford
Bradford reaktifi: Coomassie Brilliant Blue G-250 etanol içinde çözünerek hazırlanır.
(Bu çözelti hazır halde verilecektir.)
Tüp
No
Protein Stok Çöz.
(µL)
Destile
su (µL)
Bradford
Reaktifi (µL)
1
0
800
200
2
3
4
5
6
20
40
60
80
Örnek (2)
780
760
740
720
798
200
200
200
200
200
Karıştırılır ve 10 dk.
inkübe edilir
Protein stok çözeltisi: 2 mg/mL ovalbümin stok çözeltisi 0.1mg/mL olacak şekilde
seyreltilerek hazırlanır.
Absorb.
(595
nm)
Protein Kons.
(µg/mL)
VERİLERİN KULLANIMI
Bradford yöntemi kullanılarak, bir önceki denemede elde edilen tüm ekstraktların protein
konsantrasyonları tayin edilir ve µg/mL cinsinden aşağıdaki tabloya kaydedilir.
Karaciğer
%20 Tuz
Karaciğer
%80 Tuz
Yumurta
Sarısı
%50 Tuz
Yumurta Akı
Aseton
Yumurta
Sarısı Aseton
Karaciğer
Aseton
Yumurta
Akı
%50 Tuz
1. Grup
2. Grup
3. Grup
1. Grup
2. Grup
3. Grup
ÇALIŞMA SORULARI
1- Protein kimyasında Coomassie boyalarının kullanım alanları nelerdir?
Biyokimya Laboratuvarı I – Güz
44
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY VIII
PROTEİNLERİN ELEKTROFOREZ İLE AYRILMASI VE
MOLEKÜL AĞIRLIKLARININ TAYİNİ
Elektroforez, bir elektrik alan altındaki moleküllerin, bu alanda farklı hareket etmeleri
esasına dayanılarak yapılan bir ayırma yöntemidir. Bu elektrik alan altında pozitif yük
taşıyan proteinler katoda, negatif yük taşıyan proteinler anoda doğru göç ederler. Bu
göçün hızı çeşitli faktörlerden etkilenir;
a- Yükün büyüklüğü: Net yük arttıkça proteinlerin göç hızları da artar.
b- Molekül büyüklüğü: Proteinin büyüklüğü arttıkça göç hızı azalır. Burada
proteinin molekül ağırlığı olduğu kadar molekülün şekli de önemlidir.
c- Destek ortamının özelliği: Destek ortamındaki gözeneklerin boyutu küçüldükçe,
proteinin göç hızı azalır.
d- Elektrik alanın büyüklüğü: Voltaj arttıkça göç hızı da artar.
e- Tamponun iyonik gücü: Tamponun iyonik gücü azaldıkça göç hızı artar.
f- Sıcaklık: Sıcaklık arttıkça göç hızı artar.
Elektroforez günümüzde saflaştırılmış bir protein örneğinin saflığının kontrolünde, küçük
miktarlarda proteinin elde edilmesinde, bir örnekteki proteinlerin kalitatif ve kantitatif
analizlerinde kullanılmaktadır. Kâğıt elektroforezi, bölge elektroforezi, SDSPoliakrilamid jel elektroforezi (SDS-PAGE), immüno elektroforez, DNA elektroforezi
gibi çok çeşitli elektroforez yöntemleri mevcuttur.
Günümüzde proteinlerin analitik tayinleri için en çok kullanılan yöntem poliakrilamid
jel elektroforezidir. Bu yöntemde destek maddesi olarak poliakrilamid polimerinden
yararlanılır. Poliakrilamid jel uzun polimer zincirleri oluşturan, aktive edilmiş akrilamid
moleküllerinin reaksiyonu ile oluşur. Çözeltide bu zincirler tek başlarına birleşemezler.
Jelin oluşumunda zincirlerin çapraz bağlanması için N,N′-metilenbisakrilamid
kullanılır.
Şekil 8.1- Poliakrilamid polimeri
Biyokimya Laboratuvarı I – Güz
45
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Bu elektroforez yöntemi de çeşitli alt gruplara ayrılır. PAGE’de yatay, dikey kasetler ve
dikey diskler kullanılabilir. Kasetler farklı boyutlarda olabilir. Kaset boyutu
büyüdüğünde yapılan ayırmanın çözünürlüğü ve gücü artar ancak işlem süresi uzar ve
jelin çalışılması zorlaşır.
a- Doğal şartlarda PAGE: Ayırma analizlenen proteinin yük ve boyutuna göre
yapılır. Genellikle alkali (pH 8–9) tampon sistemleri kullanılır. Proteinler
denatüre olmadan yürütüldükleri için elektroforez sonunda proteinler
aktivitelerini kaybetmezler ve geri kazanılabilirler. Ancak bu sistem sadece
çözünür proteinler ve elektroforez sırasında agregat oluşturmayan proteinler için
kullanılabilir.
b- Denatüre edici şartlar altında SDS-PAGE: SDS anyonik bir deterjandır. Negatif
yüklü SDS molekülleri protein molekülünü kaplar ve proteinlerin net yükünü
maskeler. Tüm proteinler negatif yükle kaplanmış olduklarından ayırma yüke
göre değil sadece molekül boyutuna bağlıdır.
Şekil 8.2- SDS ile proteinlerin negatif yüklenmesi
SDS-PAGE’e
uygulanacak
olan
örnekler,
merkaptoetanol, SDS, bromfenol mavisi ve gliserol
içeren tamponla muamele edilirler. Merkaptoetanol
proteinin tersiyer yapısını bir arada tutan disülfit
bağlarını indirger. SDS proteini denatüre ederek doğal
yapısını bozar ve molekülü eksi yüklü bir kılıfla
kaplar. Farklı konsantrasyonlarda (%7.5, %10, %12.5
akrilamid/bisakrilamid konsantrasyonu içeren) jeller
kullanılarak elektroforez gerçekleştirilebilir. Düşük
konsantrasyonlu jeller büyük molekül ağırlıklı
proteinlerin, yüksek konsantrasyonlu jeller ise küçük
molekül ağırlıklı proteinleri ayırmak için kullanılır.
c- İki boyutlu PAGE: İki boyutlu poliakrilamid jel
elektroforezinde proteinler izoelektrik odaklama (IEF)
ve SDS-PAGE yöntemleri kullanılarak ayrılırlar. Bu
yöntemlerin her biri jel başına 100 bileşeni ayırma
gücüne sahipken, birleştiklerinde yaklaşık 10000
bileşenin ayrılmasını sağlarlar. IEF yönteminde pH
gradyanı oluşturan çeşitli amfolitler kullanılarak
proteinler önce izoelektrik noktalarına göre ayrılır.
Daha sonra aynı proteinler SDS-PAGE yöntemi
kullanılarak molekül büyüklüklerine göre ayrılırlar.
Biyokimya Laboratuvarı I – Güz
Şekil 8.3- İki boyutlu jel elektroforezi
46
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 8.4- İki boyutlu elektroforez ile elde edilmiş jel görüntüsü
Şekilde iki boyutlu poliakrilamid jel elektroforezi sonucunda elde edilen bir jel
gösterilmektedir. Buradaki her bir nokta bir protein veya polipeptide karşılık gelir.
KİMYASALLAR
Amonyumperoksodisülfat, akrilamid, bisakrilamid, tris, TEMED, SDS, glisin, HCl, βmerkaptoetanol, bromfenol mavisi, metanol, asetik asit, Coomassie G250, fosforik asit,
amonyum sülfat.
YÖNTEM
Çözeltilerin hazırlanması:
Amonyumperoksodisülfat (APS): %10 (w/v) oranında taze olarak hazırlanır.
Akrilamid/Bisakrilamid çözeltisi: 29.2 g akrilamid ve 0.9 g bisakrilamid destile su içinde
çözülerek hacmi 100 mL’ye tamamlanır ve +4oC’de saklanır.
1.5 M Tris-HCl çözeltisi (pH 8.8): 18.15 g tris baz tartılır ve bir miktar destile suda
çözüldükten sonra pH’sı 6 N HCl ile 8.8’e ayarlanır ve son hacmi 100 mL’ye
tamamlanarak +4 oC’de saklanır.
1 M Tris-HCl çözeltisi (pH 6.8): 12.11 g tris baz tartılır ve bir miktar destile suda
çözüldükten sonra pH’sı 6 N HCl ile 6.8’e ayarlanır ve son hacmi 100 mL’ye
tamamlanarak +4 oC’de saklanır.
SDS çözeltisi: %10 (w/v) oranında taze olarak hazırlanır.
PAGE Tamponu: 3 g Tris baz, 15 g Glisin ve 1 g SDS bir miktar destile suda çözüldükten
sonra 1 L’ye hacmi tamamlanır (her elektroforez işleminde taze hazırlanır).
Biyokimya Laboratuvarı I – Güz
47
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
SDS-PAGE jel kasetinin hazırlanması: İçinde jelin polimerize olacağı kuru ve temiz
cam levhalar aralarına plastik ayırıcı konularak üst üste konulur. Kıskaçlar takılarak
döküm standına yerleştirilir.
Şekil 8.5- Jel kasetlerinin hazırlanması
Poliakrilamid jelin hazırlanması: Aşağıdaki tabloya göre jel çözeltisi hazırlanır.
65 mL %12.5’lik Alt Jel Kompozisyonu
Akrilamid/Bisakrilamid
26 mL
Tris-HCl 1.5 M pH 8.8
16.25 mL
SDS %10 (w/v)
650 µL
Destile su
21,64 mL
APS %10 (w/v)
350 µL
TEMED
70 µL
20 mL %4.5’lik Üst Jel Kompozisyonu
Akrilamid/Bisakrilamid
3 mL
Tris-HCl 1 M pH 6.8
2.5 mL
SDS %10 (w/v)
200 µL
Destile su
15 mL
APS %10 (w/v)
70 µL
TEMED
20 µL
Biyokimya Laboratuvarı I – Güz
48
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
TEMED ve APS çözeltileri polimerleşmeyi başlatacak olan çözeltiler oldukları için
jel çözeltisinin kasete dökülmesinden hemen önce ilave edilir. Hazırlanan alt jel
çözeltisi iyice karıştırılır ve jel kasetlerinin içine kasetin 4/5’i dolacak şekilde dikkatle
dökülür. Üzerine bir miktar n-butanol çözeltisi ilave edilir ve polimerleşmenin
tamamlanması beklenir.
Polimerleşme tamamlandıktan sonra n-butanol çözeltisi kasetin içinden alınır.
Hazırlanan üst jel çözeltisine TEMED ve APS ilave edilerek iyice karıştırılır ve jel
kasedine dökülür. Döküldükten hemen sonra örneklerin tatbik edileceği kuyuların
oluşması için tarak yerleştirilir ve polimerleşme tamamlanıncaya kadar beklenir.
Polimerleşme tamamlandıktan sonra tarak kuyuların bozulmamasına dikkat edilerek
çıkartılır.
Şekil 8.6- Jellere tarağın yerleştirilmesi
PAGE Yükleme Tamponu
1 M Tris-HCl (pH 6.8)
1.0 mL
Gliserin
1.6 mL
%10 SDS (w/v)
0.2 mL
β-merkaptoetanol
0.4 mL
Destile su
8.0 mL
Bromfenol mavisi
Eser miktarda
Biyokimya Laboratuvarı I – Güz
49
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Örneklerin yüklenmesi: Protein örneklerinin
konsantrasyonları 3 µg/µL olacak şekilde hesaplanır
ve gerekli seyreltmeler PAGE yükleme tamponu ile
yapılır. Örneklerden 100 µL alınarak ependorf
tüplere konularak 1 dakika 100°C’lık su
banyosunda ısıtılarak denatüre edilirler.
Soğutulduktan sonra bir otomatik pipet yardımıyla
her kuyuya aşağıdaki şekilde gösterildiği gibi 10 µL
(kuyu başına 30 µg protein düşecek şekilde) protein
tatbik edilir.
Şekil 8.7- Örneklerin yüklenmesi
Kuyu
Numarası
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Yüklenen Protein
Karaciğer %20 Tuz
Karaciğer %80 Tuz
Yumurta Akı %50 Tuz
Yumurta Sarısı %50 Tuz
Protein Standardı
Karaciğer Aseton Grup 1
Karaciğer Aseton Grup 2
Karaciğer Aseton Grup 3
Yumurta Akı Aseton Grup 1
Yumurta Akı Aseton Grup 2
Yumurta Akı Aseton Grup 3
Protein Standardı
Yumurta Sarısı Aseton Grup 1
Yumurta Sarısı Aseton Grup 2
Yumurta Sarısı Aseton Grup 3
Şekil 8.8- Jel üzerindeki kuyuların yerleşimi
Biyokimya Laboratuvarı I – Güz
50
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Standart Protein
Sığır Albumin
Yumurta Albumin
Gliseraldehit-3-p-Dehidrojenaz
Sığır Karbonik Anhidraz
Tripsinojen
Tripsin İnhibitör
α-Laktalbumin
Molekül Ağırlığı (kDa)
66
45
36
29
24
20
14.2
Jellerin Elektroforez Cihazına Yerleştirilmeleri: Protein tatbik edilen kuyuların üzeri
SDS-PAGE yürütme tamponu ile doldurulur ve kaset aşağıdaki şekilde görüldüğü gibi
merkezi soğutma aparatına yerleştirilir. Elektroforez sırasında bu aparatın içinden geçen
su jelin fazla ısınmasını önleyerek yürümenin düzgün olmasını sağlar ve jeller için destek
görevi görür. Ayrıca jel kasetleri soğutucu aparata yerleştirildiklerinde üst tampon
haznesini oluşturur.
Şekil 8.9- Jel kasetlerinin tampon haznesine yerleştirilmesi.
Merkezi soğutma aparatı tampon haznesine yerleştirilir.
PAGE Yürütme Tamponu (1.5 L için)
Tris-Baz
Glisin
%10 SDS (w/v)
4.5 g
22.5 g
1,5 mL
Taze olarak hazırlanmış 1500 mL SDS-PAGE yürütme tamponu, öncelikle üst tampon
haznesi dolacak şekilde ve platin elektrotların ıslanmamasına dikkat edilerek tampon
haznesine doldurulur. Üst tampon haznesi dolduktan sonra PAGE yürütme tamponun
kalan kısmı tampon haznesinin altında jeller ile temas edecek seviyeye kadar yükselir ve
alt tampon haznesini oluşturur. Platin elektrotların kuruluğu kontrol edildikten sonra
doğru kutupların üst üste gelmesine dikkat edilerek güç kablolarını taşıyan kapak
kapatılır. (Kapaktaki + ucun ( kırmızı) soğutma aparatındaki + uca (kırmızı) ve – ucun
(siyah) soğutma aparatındaki – uca (siyah) denk gelmesine dikkat edilir).
Biyokimya Laboratuvarı I – Güz
51
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 8.10- Elektroforezde kullanılan güç kaynağı ve elektroforez aparatı
Jellerin yürütülmesi: Soğutma aparatının su bağlantıları takılır ve elektroforez sistemi
bir güç kaynağına bağlanır. Jel başına önce 20 mA, daha sonra 30 mA verilecek şekilde
güç kaynağı ayarlanır ve elektroforez işlemine başlanır. Bu işlemin süresi ortam
sıcaklığına, jelin konsantrasyonuna ve boyutuna bağlıdır. Bizim deneyi
gerçekleştirdiğimiz şartlarda jelin tam olarak yürümesi yaklaşık 5 saat sürer.
Elektroforez işlemi tamamlandıktan sonra jel kasedi soğutma aparatından ayrılır, plastik
ayırıcılar çıkarılır ve cam levhalar açılarak jel içinde 500 mL fiksasyon çözeltisi bulunan
bir kaba alınır. Bu aşamada jelin kırılmaması ve parçalanmaması için çok dikkatli
olunmalıdır.
Jellerin boyanması:
Jel
Fiksasyon
Çözeltisi
%50 (v/v) Metanol
%10 (v/v)Asetik asit
Boya
Çözeltisi
% 0.065 (w/v)
Coomassie G250
%34 (v/v) Metanol
3% (v/v) fosforik asit
%17 (w/v)Amonyum
sülfat
Boya
Çıkarma
Çözeltisi
%10 (v/v) Asetik Asit
250 mL metanol ve 50 mL asetik asit çözeltisi 500 mL’lik
bir balon jojeye konulur ve destile su ile hacim
tamamlanır.
A: 55 g Amonyum sülfat /220 mL destile su.
B: 0.2 g Coomassie G250/110 mL metanol + 10 mL
fosforik asit.
Boya çözeltisi, boyamadan hemen önce, karıştırılmakta
olan B çözeltisi üzerine A çözeltisi ilave edilerek
hazırlanır.
100 mL asetik asit çözeltisi 1 L’lik balon jojeye alınır ve
destile su ile hacim tamamlanır.
Jel önce fiksasyon çözeltisi içinde bir gece boyunca bekletilir. Burada amaç proteinlerin
jele sabitlenerek boyama sırasında protein kayıplarının önlenmesi ve iyi bir boyamanın
sağlanmasıdır.
Fiksasyon çözeltisi uzaklaştırıldıktan sonra jellere boya çözeltisi ilave edilir ve 2 gün
boyunca (veya spotların görünür hale gelinceye dek) jellerin boyanması sağlanır. Bu
aşamada bir orbital çalkalayıcı kullanılarak jellerin hafifçe çalkalanması sağlanır.
Biyokimya Laboratuvarı I – Güz
52
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Böylece boya çözeltisi jeldeki protein molekülleri ile bağlanır. Ancak boya jelin protein
olmayan kısımlarına da bağlanır (spesifik olmayan bağlanma).
Boya çözeltisinden çıkarılan jeller ayrı bir kaba konulurlar ve boya çıkarma çözeltisi ile
muamele edilirler. Boya çıkarma çözeltisi jel üzerindeki spesifik olarak bağlanmamış
Coomassie boyasını uzaklaştırılarak arka planın renginin koyu maviden, açık maviye
(veya şeffafa) doğru değişmesini sağlar ve protein bantlarının daha net bir şekilde
görülmesini sağlar.
Boyamaları tamamlanmış olan jellerin görüntüleri alınır ve daha sonraki işlemlerde
kullanılmak üzere saklanırlar.
Şekil 8.11- Boyama işlemi sonucu elde edilen jel görüntüsü
VERİLERİN KULLANIMI
5 ve 12 numaralı kuyulara tatbik edilmiş
olan protein standardı örneklerine ait
protein bandlarının her biri için Rf
değerleri hesaplanır. Rf değerleri x
eksenine ve molekül ağırlıklarının ln
değeri y eksenine gelecek şekilde bir
kalibrasyon grafiği çizilir. Bu grafikten
yararlanılarak diğer protein örneklerindeki
belirgin protein bantlarının hangi molekül
ağırlığına karşı geldiği hesaplanır.
Şekil 8.12- Proteinlerin molekül ağırlığının bulunması
ÇALIŞMA SORULARI
1- Proteomik (proteomics) nedir? Araştırınız.
Biyokimya Laboratuvarı I – Güz
53
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY IX
PROTEİNLERİN N- TERMİNAL AMİNOASİTLERİNİN İNCE
TABAKA KROMATOGRAFİSİ İLE BELİRLENMESİ
Proteinlerin fonksiyonları amino asit dizilimlerine bağlıdır. Çünkü bir proteinin
aminoasit dizilimi (sekuansı) o proteinin hangi kimyasal özelliklere sahip olacağını ve 3
boyutlu yapısının nasıl olacağını belirler. Bu sebeplerden dolayı proteinlerin
tanımlanmasında dizi analizlerinin yapılması gerekir. Dizi analizleri farklı yöntemlerle
gerçekleştirilebilir. Bu yöntemlerden biri Edman degradasyonudur. Aşağıda bir dizi
analizi şematik olarak gösterilmiştir.
Şekil 9.1- Proteinlerin dizi analizi
Biyokimya Laboratuvarı I – Güz
54
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Genel olarak bir dizi analizi çalışmasında şu basamaklar uygulanır.
1- Protein birden fazla alt birim içeriyorsa önce
alt birimlere ayrılır. Protein ve alt birimler
saflaştırılır.
2- Proteinde bulunan peptid içi (intra) ve
peptidler arası (inter) disülfit bağları 2merkaptoetanol, ditiyotreitol (DTT) gibi bir
ajanla indirgenir ve ortaya çıkan serbest –SH
gruplarının
sonraki
basamaktaki
asit
hidrolizinde oksitlenmelerini engellemek için
sisteinler iyodoasetat ile S-karboksimetilsistein’e dönüştürülür.
Bu aşamadan sonra örnek birkaç kısma ayrılır ve
devam eden işlemler ayrı ayrı bu protein kısımlarına
uygulanırlar.
3- Protein 6N HCl ile sıcakta muamele edilerek
amino asitlerine ayrılır ve kromatografik
yöntemlerle her bir amino asitten ne miktarda
içerdiği bulunur, yani toplam amino asit
bileşimi tayin edilir.
4- Karboksil ve amino terminal analizleri
gerçekleştirilir.
5- Proteaz enzimleri kullanılarak protein belirli
bölgelerden kırılır ve Edman degradasyonu
uygulanarak her bir parçanın amino asit
dizilimi bulunur. (Edman yöntemi sadece 5060 aminoasitten oluşan parçaların tayinine izin
verir, daha uzun proteinlerde hata oranı
artmaya başlar)
6- Bu parçaların dizilimi, amino asit bileşimi, Nve C- terminal analiz bilgileri bir araya
getirilerek proteinin amino asit dizilimi
bulunur.
Şekil 9.2- Sığır insülin proteinin amino asit dizilimi
Biyokimya Laboratuvarı I – Güz
55
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 9.3- Disülfit bağının indirgenmesi ve oksidasyonu
Yukarıda anlatıldığı gibi proteinlerin parçalara ayrılmaları için proteaz adı verilen
enzimler kullanılır. Bazı proteazlar peptid bağını sadece belli bir amino asitten sonra
(veya önce) ve bu sayede tahmin edilebilir bir şekilde kırarlar.
Proteaz veya Kimyasal Ajan
Kırılan Peptid Bağı
Tripsin
Lizin, Arjinin (C)
Kimotripsin
Fenilalanin, Triptofan, Tirozin (C)
Pepsin
Fenilalanin, Triptofan, Tirozin (N)
Termolizin
Fenilalanin, Triptofan, Tirozin (C)
Submaxillarus proteaz
Arjinin (C)
Endoproteinaz Lys C
Lizin (C)
Siyonejen Bromür
Metiyonin (C)
Karboksipeptidaz A
C Terminal Aminoasit (N)
(C): C- Terminalinden kırma yapar.
(N): N- Terminalinden kırma yapar.
Biyokimya Laboratuvarı I – Güz
56
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Örneğin;
glisinilfenilalaninilalanin
tripeptidi
kimotripsin ile muamele edilirse glisinilfenilalanin
ve alanin elde edilir. Eğer pepsin ile muamele
edilirse glisin ve fenilalaninilalanin elde edilir.
Proteinin C- terminalinin bulunması için, protein
karboksipeptidaz A ile muamele edilir ve kopan
amino asit kalıntısı tayin edilir. Ancak enzim
sürekli olarak C-terminalindeki amino asitleri
koparmaya devam edeceğinden reaksiyonun
istenilen basamakta kesilmesi önemlidir.
Şekil 9.4- Glisinilfenilalaninilalanin tripeptidi
Şekil 9.5- Karboksipeptidaz A enziminin mekanizması
Proteinin N- terminal analizi için Sanger yöntemi kullanılır. Bu yöntemde 2,4dinitroflorobenzen reaktifi kullanılarak proteinin N-terminal ucundaki amino asit 2,4dinitrofenil türevine dönüştürülür. Daha sonra asit hidrolizi ile protein amino asitlerine
ayrılır ve bir amino asit karışımı oluşur. Bu karışımdaki N-terminal amino asidinin 2,4dinitrofenil türevi kromatografik yöntemler kullanılarak tanımlanır. Ancak protein
tamamen amino asitlerine parçalandığından dizi analizinde kullanılamaz.
Proteinlerin amino asit dizi analizlerinin tayini için Edman yöntemi kullanılır. Bu
yöntemde N- terminal amino asidi fenilizotiyosiyanat ile reaksiyona sokulur. Bu amino
asit feniltiyohidantoin oluşturarak peptid zincirinden ayrılır ve kromatografik yöntemlerle
tayin edilir. Her seferinde sadece N- terminalindeki amino asit ayrıldığı ve zincirin geri
kalan kısmı korunduğu için bu işlem tekrar tekrar yapılarak zincirdeki bütün amino asitler
tanımlanabilir.
Edman
yöntemi
tamamen
otomatik
cihazlar
yardımıyla
gerçekleştirilebilir. Ancak her bir döngüde bazı yan ürünler açığa çıktığından 50-60
amino asit sonrasında tayinde hatalar artar. Bu nedenle proteinlerin daha önceden
proteazlarla parçalanması gerekir.
Biyokimya Laboratuvarı I – Güz
57
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 9.6- Edman ve Sanger yöntemi
Günümüzde elektroforez ile saflaştırılmış proteinlerin amino asit diziliminin
bulunmasında MALDI-TOF yöntemi ve protein veri bankaları kullanılmaktadır. Bu
yöntemde proteinler tripsin kullanılarak parçalara ayrılır ve kütle spektrofotometreleri ile
her bir parçanın kütle analiz şablonları çıkarılır. Bu şablonlar internet üzerindeki veri
tabanlarındaki proteinlere ait şablonlar ile karşılaştırılarak bilinmeyen proteinin ne
olduğu ve dolayısıyla amino asit dizilimi bulunabilir. Ancak bu yöntem dolaylı bir analiz
yöntemidir, daha önceden yapısı aydınlatılmamış bir proteinin amino asit diziliminin
bulunması için kullanılamaz.
Saflaştırılmış ve amino asit dizilimi belirlenmiş olan polipeptidin 3 boyutlu yapısı xışınları kristalografisi ve NMR teknikleri kullanılarak aydınlatılır. Böylece protein
saflaştırılması ve karakterizasyonu tamamlanmış olur.
Biyokimya Laboratuvarı I – Güz
58
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
KİMYASALLAR
Aspartik asit, lösin, valin, arjinin, prolin, sistein, sistin, glisin, Na2CO3, 2,4dinitroflorobenzen, HCl, kloroform, benzil alkol, glasiyel asetik asit.
YÖNTEM
50 mL’lik bir erlen içinde 20 mg amino asit %5’lik 10 mL Na2CO3 çözeltisinde çözülür.
Üzerine 0.5 mL 2,4-dinitroflorobenzen (DNFB) ilave edilir. DNFB bazı insanlarda
alerjik reaksiyonlara sebep olabileceğinden çalışma sırasında eldiven takılmalı ve
dikkatli olunmalıdır. Karışım 40°C sıcaklıkta 30 dakika devamlı karıştırılarak tutulur.
Sıcaklığın 40 °C’nin üzerine çıkmamasına dikkat edilir. Isıtma sonunda çözeltinin rengi
turuncuya döner. Reaksiyona girmemiş olan DNFB bir pipet yardımıyla uzaklaştırılır.
Daha sonra 1.5 mL derişik HCl dikkatle ilave edilir. Türev bu aşamada çöker. Eğer
çökelme olmazsa erlen buz banyosunda soğumaya bırakılır. Çökelti bir test tüpüne
alınarak az miktarda aseton içinde çözülür.
Mobil faz olarak (kloroform: benzil alkol: glasiyel asetik asit) ; (45:20:1) karışımı
kullanılır. Mobil faz 1 cm yüksekliğinde olacak şekilde kromatografi tankına konulur.
Bir TLC (thin layer chromatography; ince tabaka kromatografisi) plakasına kurşun
kalemle plakayı zedelememeye dikkat ederek alttan 3 cm yükseklikte bir çizgi çekilir. 2
cm aralıkla bu çizgi üzerinde tatbik noktaları işaretlenir. Hazırlanmış olan amino asit
türevleri bu noktalara tatbik edilir. Bütün örnekler tatbik edildikten sonra TLC plakasının
kuruması beklenir ve kromatografi tankına yerleştirilir. Mobil fazın plaka üzerinde üstten
2 cm kalıncaya kadar yürümesi beklenir. Plaka tanktan çıkarılır, mobil fazın yürüme
sınırı kurşun kalem ile işaretlenir. Çeker ocakta kurutulur. Her bir aminoasidin ve
türevinin Rf değerleri hesaplanır.
ÇALIŞMA SORULARI
1- Proteinlerin 3 boyutlu yapı aydınlatılması nasıl yapılır? Araştırınız.
Biyokimya Laboratuvarı I – Güz
59
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DENEY X
MEMELİ DOKULARINDAN DNA İZOLASYONU VE
BAZI ÖZELLİKLERİNİN İNCELENMESİ
DNA ve RNA nükleik asitler olarak adlandırılırlar ve nesilden nesile kalıtımsal bilginin
aktarılmasını sağlayan uzun zincirli polimerlerdir. Bu makromoleküllerin yapı taşları
nükleotidlerdir. Her nükleotid bir şeker, bir fosfat ve bir bazdan oluşur.
Şekil 10.1- DNA ve RNA yapısında yer alan azotlu bazların yapıları
Şekil 10.2- Deaoksinükleotid yapısı
Nükleik asitler nükleotidlerin 3′, 5′-fosfodiester köprüleri ile bir araya gelmesinden
oluşmuştur. Bu zincirlerde fosfodiester köprüleri ile bağlanmış şeker molekülleri nükleik
asitlerin iskeletini oluşturur.
Biyokimya Laboratuvarı I – Güz
60
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Şekil 10.3- Nükleik asit yapısı
Şekil 10.4- DNA yapısında yer alan baz çiftleri
Biyokimya Laboratuvarı I – Güz
61
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DNA ikili sarmalında adenin-timin (A-T) ve guanin-sitozin (G-C) bağları kurulur. DNA
ikili sarmalını bir arada tutan kuvvetler, nükleotid zincirleri arasındaki A-T ve G-C
çiftleri arasındaki hidrojen köprüleri (yatay interaksiyon) ve nükleotid zincirleri
üzerindeki bazlar arasındaki etkileşimlerdir (dikey interaksiyon).
Şekil 10.5- DNA çift sarmal yapısı
Protein molekülünde olduğu gibi DNA
molekülünün
yapısındaki
herhangi
bir
bozulmaya da denatürasyon denir. DNA
denatürasyonu yatay ve dikey çekici güçlerden
herhangi birindeki azalma veya kopma sonucu
olur. Yatay güçlerdeki azalma ya da kopma
sonucu olan denatürasyon DNA’nın 260
nm’deki
absorbansının
artması
ile
belirlenebilir. Dikey güçlerdeki azalma ya da
kopma
sonucu
olan
denatürasyon
(hipokromisite) tek başına 260 nm’de
absorbans değişikliğine sebep olmaz. Ancak
DNA’nın 210 nm civarındaki 2. pikinin
absorbansında azalma hatta tamamen yok olma
ve pikin dalga boyunun daha yüksek dalga
boylarına kayması meydana gelir.
DNA sarmalını oluşturan zincirlerin birindeki veya her ikisindeki bazlardan bir veya bir
kaçının kopması sonucu nükleotid zincirde kısalma meydana geliyorsa bu olaya
DNA’nın degradasyonu denir. Degradasyon gerçekleştiğinde 260 nm’deki absorbansta
azalma meydana gelir.
42°C’ın üzerindeki sıcaklıklarda DNA ikili sarmalı
arasındaki hidrojen köprüleri öncelikle A-T zengin
bölgelerde olmak üzere gevşeyip açılmaya başlar.
Kullanılan tampon çözeltiye bağlı olarak 56-80°C
arasındaki sıcaklıklarda DNA ikili sarmalı kendisini
oluşturan nükleotid zincirlerine ayrılır. Buna DNA’nın
erimesi denir. DNA heliksinin %50’sinin denatüre
olduğu sıcaklığa DNA’nın erime sıcaklığı (Tm) adı
verilir. Tm değeri o DNA’nın A-T/G-C oranına bağlı
olduğundan, DNA’nın tanımlanmasını sağlayan
spesifik bir değerdir (yani bir fareden alınan DNA’nın
Tm değeri, bir insandan alınan DNA’nın Tm değerinden
farklı olacaktır veya aynı bakterinin iki farklı suşunun
Şekil 10.6- DNA‘nın erimesi
DNA’larının Tm değerleri farklı olacaktır). Ayrıca Tm
değeri kullanılan tampon çözeltinin iyonik şiddetine ve pH’sına da bağlıdır. Bu nedenle
Tm değeri verilirken mutlaka deneyin hangi, şartlarda yapıldığı da belirtilmelidir.
Biyokimya Laboratuvarı I – Güz
62
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DNA erimesi sıcaklık 90-100 °C’nin üzerine çıkılmadığı sürece geri dönüşümlüdür. Eğer
80°C’ye ısıtılmış bir DNA çözeltisi, oda sıcaklığında yavaş yavaş soğumaya bırakılırsa
iki sarmal tekrar birleşerek DNA ikili sarmalını oluştururlar (hibridizasyon). Ancak aynı
DNA oda sıcaklığında değil de bir buz banyosunda ani soğutmaya bırakılacak olursa
DNA sarmalarında hibridizasyon gerçekleşmez ve nükleotid zincirleri birleşmezler.
Ayrıca DNA boyunda kısalmalar, yani DNA degradasyonu meydana gelebilir. Eğer
degradasyon meydana gelmediyse bu çözeltiler yeniden ısıtılıp yavaş yavaş soğumaya
bırakılırlarsa DNA ikili sarmalını yeniden oluşturabilirler.
Şekil 10.7- DNA’nın erimesine G-C içeriğinin etkisi
Yukarıda anlatıldığı gibi nükleik asitler 260 nm dalga boyunda absorbans piki verirler.
Bu absorbans değeri kullanılarak nükleik asidin konsantrasyonu tayin edilebilir. 260 nm
dalga boyunda, 1 cm’lik kuartz küvetlerde 1’lik bir absorbans 50µg/mL DNA
konsantrasyonuna karşılık gelir. Tek zincirli DNA veya RNA içinse bu değer 40
µg/mL’dir.
Nükleik asitlerin saflığını tayin etmek için nükleik asit çözeltilerinin absorbanslarının
240-300 nm arasında ölçülmesi gerekir. Proteinler 280 nm’de pik verdiklerinden A260 nm/
A280 nm oranından yararlanılarak saflık tayini yapılabilir. DNA için bu oran 1.8, RNA
içinse 2.0 olmalıdır. Eğer bu verilen değerlerden daha düşük değerler elde ediliyorsa, bu
örneğin proteinlerle kontamine olduğunu gösterir. Kontaminasyon bir kimyasal karışım
içinde düşük miktarlarda (genellikle istenmeyen) başka maddelerin var olmasıdır.
Hücreler (hücre tiplerine göre farklılık göstermekle birlikte) büyük ölçüde su
içerdiklerinden, hücre organellerinin yapısal özellikleri su ile reaksiyonları sonucu ortaya
çıkar. Hücreler ve hücre organelleri yapısal özellik ve bütünlüklerini izotonik çözeltiler
içinde koruyabilirler. Hücrelerle aynı osmatik basınca sahip çözeltilere izotonik
çözeltiler denir. İzotonik çözeltilerde hücre ile etrafındaki çözelti arasındaki su alış verişi
dengededir. %0.876’lık NaCl (serum fizyolojik), %5 dekstroz (serumda kullanılır) gibi
çözeltiler yaygın olarak kullanılan izotonik çözeltilere örnek olarak verilebilirler.
Biyokimya Laboratuvarı I – Güz
63
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
Her ne kadar hemen hemen tüm hücreler DNA içerseler de bazı dokularda bu miktar çok
az olduğundan bu tip dokular DNA izole etmek için iyi bir kaynak değildirler. Bazı
dokular ise yüksek deoksiribonükleaz aktivitesine sahip olduklarından DNA izolasyon
esnasında küçük parçalara ayrılır. Bu nedenle DNA elde etmek için yüksek DNA ve
düşük nükleaz aktivitesine sahip dokular tercih edilmelidir. Bu amaçla en iyi kaynaklar
timus ve lenf en iyi dokulardır. Dalak, testis ve yumurtalıklarda DNA elde etmek için
kullanılabilirler.
KİMYASALLAR
NaCl, EDTA, SDS, etanol.
YÖNTEM
Çözeltilerin hazırlanması:
Ekstraksiyon çözeltisi: 0.15 M NaCl ve 0.1 M EDTA çözeltisinden oluşur. 0,925 g
EDTA ve 2,19 g NaCl 250 mL destile su içinde çözülerek hazırlanır. Deneyde soğuk
olarak kullanılır.
2M NaCl çözeltisi: 11.7 g NaCl 100 mL destile suda çözülerek hazırlanır.
DNA’nın İzolasyonu:
DNA izolasyonunun yapılacağı dokudan yaklaşık 10 g alınır ve 100 mL ekstraksiyon
çözeltisi ilave edilir. Daha sonra kısa sürelerle ve DNA içeren çözeltinin ısınmamasına
dikkat edilerek homojenizatörde homojenize edilir. DNA’nın bozulmaması için bu
işlemler soğukta yapılmalı ve kullanılan çözeltiler önceden soğutulmuş olmalıdır.
Homojenize hale getirilmiş olan çözeltiye 1 mL %10 SDS ilave edilir. Hafifçe alt üst
edilerek çözelti karıştırılır (SDS deterjan olduğundan köpürmemesine dikkat edilir). Daha
sonra 4000 rpm’de 5 dakika santrifüj edilir. Üstteki çözelti (süpernatant) dikkatle alınır
ve temiz bir santrifüj tüpüne hacim 4 mL olacak şekilde aktarılır.
Süpernatant çözeltisine 8 mL soğuk 2M NaCl çözeltisi ilave edilir ve dikkatle karıştırılır.
Daha sonra 4000 rpm’de 5 dakika santrifüj edilir. Süpernatant yaklaşık 4 mL hacimde
olacak şekilde dikkatle temiz bir santrifüj tüpüne alınır.
Süpernatanta yaklaşık 8 mL soğuk etanol yavaş yavaş ilave edilir ve tüpteki değişim
gözlenir. Daha sonra 4000 rpm’de 1-2 dakika santrifüj edilir. Santrifüj sonunda üstteki
süpernatant dekante edilir ve beyaz renkli DNA presipitatı alınır. Etanol kalıntısını
uzaklaştırmak için 1-2 mL 0.1 M EDTA çözeltisi ile, karıştırılmadan kalıntılar yıkanır,
yıkama çözeltisi dekante edilerek uzaklaştırılır ve daha sonra yeterli miktarda
ekstraksiyon çözeltisinde çözülür. Çözünmeyen kısımlar santrifüj ile uzaklaştırılır. Elde
edilen DNA çözeltisi derin dondurucuda uzun süre saklanabilir.
Biyokimya Laboratuvarı I – Güz
64
Marmara Üniversitesi
Fen-Edebiyat Fakültesi
Kimya Bölümü / Biyokimya Anabilim Dalı
DNA’nın Erimesi:
DNA çözeltisinin 260 nm ve 280 nm’deki absorbansları ölçülür ve konsantrasyonu ve
saflığı tayin edilir.
4 cam tüpe elde edilen DNA çözeltisinden yaklaşık 5’er mL alınır. Tüplerden bir tanesi
oda sıcaklığında bekletilirken diğer üçü 80°C sıcaklıktaki su banyosunda 10 dakika
bekletilir. Bu sürenin sonunda bir tüp oda sıcaklığında, bir tüp buz banyosunda ve bir tüp
de –80°C soğutucuda 5 dakika tutulur. Soğutulmuş olan tüplerin oda sıcaklığına gelmesi
beklenir ve 4 tüpün absorbans değerleri 260 nm’de ekstraksiyon çözeltisi kör olarak
kullanılarak ölçülür.
Tüp Hacim
No
(mL)
Uygulanan
işlem
Oda
sıcaklığında
bekletilir.
1
5
2
5
80°C ısıtıtma
(10 dakika)
3
5
80°C ısıtıtma
(10 dakika)
4
5
80°C ısıtıtma
(10 dakika)
Uygulanan
işlem
Oda
sıcaklığında
bekletilir.
Oda
sıcaklığında
bekletilir.
Buz
Banyosunda
Bekletilir (5
dakika)
-80°C
soğutucuda
bekletilir (5
dakika)
Uygulanan
işlem
Oda
sıcaklığında
bekletilir.
Oda
sıcaklığında
bekletilir.
Oda
sıcaklığında
bekletilir.
Absorbans Absorbans
(260 nm)
(280 nm)
Ölçüm
yapılmaz
Oda
sıcaklığında
bekletilir.
ÇALIŞMA SORULARI
1- Genomik (Genomics) nedir? Genomik çalışmalar niçin önemlidir?
2- Bilimsel araştırmalarda DNA izolasyonunda genellikle fenol ekstraksiyonu
kullanılır. Bu yöntemin nasıl yapıldığını ve prensibini araştırın.
Biyokimya Laboratuvarı I – Güz
65