HÜCRE SİNYALİZASYONU
Prof.Dr.Gönül kanıgür-Sultuybek
Prof.Dr.Selma Yılmazer
• Çok hücreli organizmalarda hücreler arası
bilgi ve iletişim çok sayıda farklı molekülle
sağlanmaktadır.Bu moleküller Sinyal
molekülleri(ligand) olarak adlandırılır.
• Tek hücreli organizmalarda çevrelerinden
sinyal alırlar ve sinyale yanıt verirler.
• Örn;bakteriler
• Çok hücreli, organizmalarda hücrelerin
entegre ve koordine çalışması sinyal
molekülleri sayesindedir
• Sinyal molekülleri(ligand) uyarı oluşturmak
isteyen hücre tarafından sentezlenir ve
salgılanır.
• Salgılanan bu ligand hedef hücre zarında
,sitoplazmasında yada nükleusunda yer alan
reseptör adı verilen glikoproteine bağlanarak
hücresel cevaba yol açar.Reseptörler membrane
reseptörleri ve intrasellüler reseptörler olarak
ikiye ayrılır.
• Membran reseptörler
membrane
Glikoprotein
• Intrasellular receptors
sitozol veya nükleusda
DNAya bağlanan protein
Sinyalleyici
(ekstrasellüler )moleküller
protein & peptidler: Hormonlar,sitokinler
AA & türevleri: Gly, Glu, adrenaline,
tiroksine
Steroid: Sex Hormone,
glucocorticosteroid
yağ asidi ve türevleri: prostaglandin
Sinyalleyici( İntrasellüler)
moleküller
• Ca2+
iyonlar
• DG, ceramide
• IP3
lipid türevleri
karbohidrat
türevleri
• cAMP cGMP
nükleotidler
• Ras, JAK, Raf
proteinler
. HormonunReseptörüne bağlanma
özellikleri
• yüksek spesifiklik
• yüksek affinite
• Doyum
• Reversible bağlanma
• spesifik fonksiyon
Reseptor fonksiyonu
(1)Ligandın tanınması
(2)Liganda bağlanma
(3) Signal transdüksiyon
biological etki
Sinyalin alınması
• Ekstraselüler sinyal molekülleri hedef
hücrelerin yüzeyindeki veya içindeki özel
reseptörler tarafından tanınır.
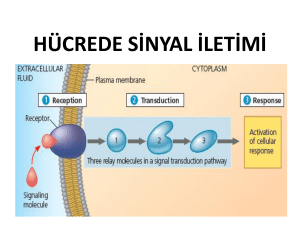
I. HÜCRE SİNYALİZASYONU
1. Hücre sinyalizasyonundaki aşamalar
Hücre
(1) (2)
(4)
(4) Sinyalleyici molekülün re
septörü tarafından tanınması
(3)
(5).intrasellüler sinyal oluşumu
Gen ekspresyon ve hücre metabolismasın
(1).sinyalleyici molekülün sentezi
.
(2sinyalleyici molekülün salgılanması
(3).sinyalleyici molekülün hedef hücreye
transportu
(6).hücresel cevabın sonlanması
(6)
(6)
Büyüme, farklılaşma, apoptosis gibi
•
•
•
•
•
•
•
Sinyalizasyon sonucu
-hücre metabolizması
-hücre hareketi
-hücre çoğalması
-hücre sağkalımı
-hücre faklılaşması
-hücre apoptozu gibi hücre davranışını
düzenleyen bir seri hücre reaksiyonu
oluşur
Salgılanan moleküllerle sinyal
iletim şekilleri:
•
•
•
•
A-Parakrin etkileri kısa sürelidir.
B-Sinaptik etkileri kısa sürelidir.
C-Endokrin etkileri uzun sürelidir.
D-Otokrin
• Bazı sinyal molekülleri sinyalleri uzun
mesafelere taşırken bazıları komşu
hücreler arasında bilgi iletirler.
• Ekstrasellüler sinyaller olarak ifade edilen
bu sinyaller etkileyeceği hedef hücrenin
uzaklığına bağlı olarak 4 farklı uyarı
mekanizması gösterirler.
Parakrin sinyal
• Sinyal molekül [lokal ] bölgesel aracılar olarak rol
oynar.
• Komşu [yakınındaki hüc] davranışını
düzenler.örneğin pancreas da
– Glukagon; somatostatin insülin salgılayan
hücreler üzerine etki eder.
Nöronal (Sinaptik) Sinyal
• Nöron hücre [sinir ] tarafından oluşan sinyalleri
verilebilir.asetilkolin ,noradrenalin.
• Nöronlar uzun aksonları ile uzaktaki hedef hüc.ler ile
temas kurarlar. Çevreden yada diğer nöronlardan gelen
sinyaller nöron aksonu boyunca elektrik impulsları
şeklinde iletilir. Bu impuls nörotransmitterlerin
salgılanmasını sağlar.
Endokrin sinyal
• Endokrin hücreler hormon sinyal mol. Kan damarı içine
salgılar. Hormon kan dolaşım yoluyla transport edilir.
Farklı hedef hücrelere ulaşan hormon reseptörü ile
etkileşerek sinyali hücre içine iletir.
• Örnek;
– Estradiol,insülin,thyroxin,adrenalin
Sinyalleyici molekül olarak
gazlar (NO,CO)
• Etkileri kısa sürelidir.NO sinir sistemi
bağışıklık sistemi ve dolaşım sistemi için
önemli bir sinyal molekülüdür.steroidler
gibi NO,CO de
• Hedef hüc plasma zarından difüzyon
yoluyla içeri girer.NO hücre içi hedef
enzimlerin aktivitesini değiştirir.yar.ömrü
birkaç saniye.komşu hüc.bölgesel olarak
etkiler..
Otokrin sinyal
• Hücrelerin kendisi tarafından oluşturulur
diger bir deyişle hücreler kendi kendilerine
sinyal gönderirler
• İmmün Sistem Hücrelerinde
– Büyüme faktörleri ,interlökinler gibi
moleküller,
– Hücre farklılaşması ve çogalması için tetik
mekanizmasıdır. Ayrıca gelişme
esnasında hücre farklılaşmasında otokrin
sinyaller kritik rol oynar.
– Otokrin sinyalindeki anormallik ise kanser
hücrelerinin çogalmasına neden olur.
• Kanser:sağ kalımı kontrol eden sinyal
yolaklarından birisinin bozulması sonucu
ortaya çıkar.
• Kanser hücrelerinin büyüme faktörlerine
gereksinim duymamalarının nedeni hücre içi sinyal
sistemlerinde gerçekleşen bozukluklardır. Hücre
çoğalmasını sağlayan sinyal yollarında görev
yapan büyüme faktörü reseptörlerinin ya da başka
proteinlerin kontrolsüz aktivitesinden kaynaklanır.
Erişkinde
otokrin sinyalleşme;
• Eicosanoid
- yağ asidi türevleri
- Arachidonic asitden yapılır
- Erişkin memelilerde otokrin sinyal
molekülleri
Otokrin sinyal molekülleri
(prostoglandinler, prostacyclin,
tromboxan,lökotrien)
Eicosanoidlerin çeşitli biyolojik
aktiviteleri
• Düz kas kontraksiyonunu etkileme
(uterus kasında doğum sırasındaprostoglandin)
• Trombosit agregasyonunu etkileme
• Ağrı, ateş
• İltihabi reaksiyon
Doku hasarı eicosanoid sentezi
(siklooksigenaz, lipoksigenaz)
Kontakt etkileşim
• Embryo
gelişimi,erişkin
dokuların
devamlılığının
sağlanmasında çok
önemlidir.
Direkt hücre-hücre sinyal iletimi
• Bir hücre komşu hücre ile doğrudan
etkileşime girer.,
• hücre-hücre
• Hücre –hücre matriks etkileşimi
sağlıyan mol. Hücre davranışını kontrol
eder.örn;integrinler kaderinler sadece
adesyon mol.degil hücre sagkalımını
vede poliferasyonu sağlıyan sinyal mol
olarakta işlev görür.
Sinyalleyici Moleküller
Sinyalin
Tipi
Endokrin
Sinyal
Molekül
Hormonlar Bölgesel
Ligandlar
Dağılım/ Uzak
Fonksiyon hedeflere
Örnek
kan akımı
yolu ile
ulaşım /
Metabolima
Adrenaline
Kortizol
Glukagon
Insulin
Parakrin
Sinyal
Nöronal
Sinyal
Kontakt
Etkileşim
Nörotranm Memran
iter
sinyal
Molekülü
İnflamasyonun Sinir
Embriyonik
Regülasyonu
sonlanmala- hücreler,
Doku Hasarı
sonrası Hücre
Proliferasyonu
Histamine
EGF
PDGF
NGF
Nitric Oxide
Rındaki
sinapslar
Acetycholine Delta
GABA
(a trans-
membrane
protein)
Gap junctionlar ile iletişim
• Sinyalin komşu hücreler arasında
ortaklaşa kullanımı
• Küçük intraselüler sinyal molekülleri
(intraselüler mediatörler)
Ca+2
siklik AMP
Sinyal moleküllerinin oluşturduğu
cevaplar
HORMONLAR VE ETKİ
MEKANİZMALARI
• Bütün biyolojik ,dolaşımdaki faktörler
tarafından kontrol altında tutulur.
• Hücre sinyalizasyonuna neden olan bu
maddeler hayvanlarda hormonlardır.
• Hormonlar büyüme gelişme ve her iki
durumda da hücreler üzerine etkindir.
• Hormonların bazıları münferit genler veya
çok sayıda gen topluluğu üzerinde direkt
etki gösterirken (steroidler),
• Bazıları (protein hormonlar) ise enzimlerin
aktivitelerini (fosforilasyon,defosforilasyon)
etkileyerek
• Hücresel cevap oluştururlar
Steroid hormonların etki
mekanizması
• Gonadlar ve adrenal bezlerden salgılanan
steroid hormonların en önemli sınıfını
androjenler,östrojenler,progestinler,glikoko
rtikoidler,mineralokortikoidler oluşur.
• Pasif difüzyonla hücreye giren
steroidler,spesifik reseptörleri aracılığıyla
,hedef hücrelerde hedef genlerin
transkripsiyonunu düzenlerler ,etkileri
nükleerdir.
• Farklı genlerin aktivasyonuyla farklı
steroidlerin farklı etkileri ortaya çıkar
• Örn;gonadal
steroidler(androjen,progesteron,östrojen)
• reprodüktif dokularda seksüel karakterler
üzerinde etkinliğe sahipken,büyümenin
düzenlenmesiyle de ilişkilidir.
• Adrenal steroidler metabolizmanın ve
,elektrolit hemoztazının kontrolünde
metabolik düzenleyiciler olarak işlev görür.
Steroid hormon reseptörlerinin hücrede
yerleşimi
steroid hormon reseptörleri sentezlendikten
sonra sitozolde kalır reseptörler
A,B,C,D,E domain’ den oluşur..
heat-shock(Şaporon) proteinleri (Hsp90,
Hsp70 ve diğerleri) ile C-terminal uçta
bulunan D/E domainleri aracılığıyla
etkileşir.
•
Ligandın reseptörüne bağlanması
takiben heat shok proteinlerden
reseptörün ayrılmasına neden
olur.hormon –Reseptör
kompleksinin,dimerizasyonu
gerçekleşir.hormon-reseptör
dimerlerinin, nukleusa transportu,
nuklear por kompleksinden
gerçekleşmesi ,reseptörün nuklear
lokalizasyon sinyali olarak iş gören D
domain’indeki temel (lizin-arginin)
kalıntılara (rezidue) bağlıdır.
1.
2.
3.
4.
5.
6.
7.
8.
Steroid taşıyıcı proteinle hücreye gelir ve taşıyıcı proteinden
ayrılır.
steroid hormon taşıyıcı proteinden ayrılır ve diffzüyon ile hücre
içine geçer.
Steroid hormonlar (Testosteron, Progesteron, Östrogen) Hsp ile
bağlı olan sitoplazmik proteine bağlanır.
Hsp ayrılır ve hormon-reseptör kompleksi oluşur yani reseptör
aktifleşir, dimerleşir.
aktif sitoplazmik reseptör aktif transportal nükleusa girer, DNA
üzerindeki HRE dizilerine homodimer olarak bağlanır.
DNA mRNA ya transkripsiyonu olur.
mRNA nükleusdan ayrılıe ve sitoplazmik ribozomlarda proteine
translasyonu olur.
yeni sentezlenen proteinler biyolojik cevabı oluşturur.
örnegin; Östrojen plazma zarından direkt
difüzyon ile geçer ve çekirdekteki
reseptörüne bağlanır.
Hormon
olmadığında,
östrojen
reseptörü Hsp90’a bağlanır. Östrojenin
bağlanması ile reseptör Hsp90’dan
ayrılır ve reseptör dimerleri oluşur.
Dimerler DNA’ya bağlanır, histon
asetiltransferaz
(HAT)
ve
koaktivatörler ile etkileşime girer ve
hedef genlerin transkripsiyonunu uyarır.
KOREGÜLATÖRLER
• Nükleer reseptörlerin hormonla bağlanması ilişkili genin
mRNA transkripsiyonun stimüle eden veya inhibe eden
farlı proteinleri etkiler. Bunlara transkripsiyon
coregülatörleri denir.
• Bu coregulatörler genlerin transkripsiyonunu aktive
ederlerse coaktivatör, inhibe ederse cosupresor denir.
•
Bu coregulatörler fonksiyonlarını diğer koregültör
proteinleri bağlayıp işlevlerini kuvvetlendirerek veya
kromatini dolayısıyla DNAyı etkileyerek transkripsiyonu
ya kolaylaştırır veya zorlaştırırlar.
Koaktivatörler
• Reseptörün agonist liganda(östrogen,testesteron gibi) bağlı olduğu
durumdaki konformasyonu koaktivatör adı verilen bazı proteinlerle
kompleks yapmasını sağlar.
•
Koaktivatörlerin intrensek bir histon asetil transferaz(HAT) aktivitesi
vardır ve bu yolla kromatin yapısındaki histonların asetillenmesi,
DNA ile histonlar arasındaki iyonik etkileşimi bozarak kompakt
kromatin yapısının gevşeyip çözülmesine sebep olmaktadır.
• Bu yapı, RNA polimeraz II’nin DNA’ya sıkıca yerleşip
transkripsiyonu başlatmasına olanak vermektedir
• HAT transkripsiyonel aktivitesi olan histon proteinlerindeki lizin
AA’ne CoAdan asetil grup ekler.
Kosupresörler
• Reseptöre antagonist bir ligand (glıkokortikoid, progestron) bağlı
olduğunda (bazı reseptörler için herhangi bir ligand bağlı olmadığı
durumda bile) ise kosupresör adı verilen proteinler reseptörle
kompleks yapar. Bu kompleksin içinde histon deasetilaz
(HDAC)enzimleri de bulunur.
• Yani DNA ile histonların ilişkisini attırarak transkripsiyonu supresse
ederler.
•
Histon deasetilazların etkisi ile asetil grupları uzaklaştırılan histonlar
tekrar pozitif yüklerini kazanır ve DNA ile sıkı etkileşime girerler.
Böylece kromatin kompakt hale gelir ve transkripsiyon önlenmiş
olur.
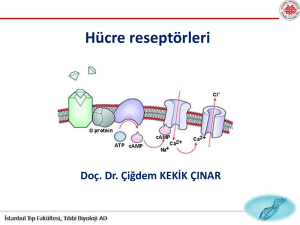
Protein hormonlar
Receptörler
– İntrasellüler
– hidrofobik(llipofilik)ligandlara baglananlar
° Nükleer hormon reseptörleri
° Guanylate siklaz(nitrikoksid receptor)
• Hücre yüzey
– hidrofilik ligandlara baglananlar
° Enzimatik ve enzime baglı reseptörler
– e.g. Reseptör tirozin kinazlar
° G-proteinleri ile baglantılı reseptörler
– a.k.a 7-transmembrane, bölgesi olan
[GPLR] reseptörler
İyon kanalına baglı olanlar
©2002 Lee Bardwell
1-Enzim ilişkili reseptörler
Reseptör protein-tirozin kinazlar
• En büyük reseptör ailesi
• Büyüme faktörü reseptörlerinin çoğu bu türden
(50 den çok).
• Hayvan hücresi çoğalma ve farklılaşmasını
kontrol eden mekanizma
• 1980 de keşfedildi
• Onkogenik protein,Rous Sarcoma virus:Src, ilk
bulunan protein tirozin kinaz
• Tek geçişli heliks transmembran
protein
• Sitozolik bölge protein tirozin kinaz
etkinliği gösterir
• Reseptör protein tirozin kinazlar
substratlarını tirozin kalıntılarında
fosforiller
• Çoğu tek bir polipeptid
• İnsülin reseptörü dimer
Etki mekanizması
• Ligand (GF) bağlanır
• Reseptör dimerizasyonu.
PDGF, NGF dimer yapıdadır.
Bağlanınca iki reseptör bir
araya gelir; dimer oluşur.
• Reseptörün otofosforilasyonu:
1. Protein kinaz aktivitesi artar
2. Katalitik bölge dışındaki
fosforillenme ile diğer
proteinlerin bağlanma bölgeleri
oluşur
• SH2 (Src homoloji 2) bölgesi içeren
proteinlerin bağlanması
• SH2 içeren protein membranlarda
yerleşir
• Diğer proteinlerle etkileşir
Protein hormon reseptör familyası
• Tirozin kinaz reseptör
familyası.insülin,IGF-l
gibi.bu reseptörlerin
tirozin kinaz aktivitesi
bulunur.sinyal iletimi
substrat
moleküllerininfosforilasyo
nu ile gerçekleşir.
• Sitokin .,hemopoetin
reseptör familyası.ligand
baglama bölgeleri
benzerdir.GH,prolaktin
• Üçüncü grup.7transmembran sahasına
sahip G-Proteinleri ile
baglantılı [GPLR]
reseptörlerparatiroid
hormon,LH,FSH,TSH
(3) G Protein- ilişkili Reseptörler
(serpentine R)
1) Zarı 7-kez geçen alfa heliks
yapısındadır.
2-G proteini ile ilişkili reseptörler
• En büyük reseptör ailesi
• Etkisini guanin nükleotidlerini bağlayan
proteinler (G proteinleri) aracılığı ile yapar
• 1000 den fazla çeşit GPLR reseptör proteini
tanımlandı
-nörotransmitter reseptörleri
-nöropeptid reseptörleri
-peptid hormon reseptörleri
-koku, görme, tat reseptörleri
•
•
•
•
•
Ligand
Reseptörler
G-proteinleri
Protein Kinaz and Fosfatazlar
İkincil Aracılar (C-AMP, C-GMP CaKalmodulin, Fosfotidil İnositol)
• İkincil Aracıların substratları
©2002 Lee Bardwell
G-proteinleri
- G-Proteinleri, reseptörle[GPLR] efektör
arasında aracılık eder. Heterotrimerik
yapıda, b, g;
- alt birimiGTP yi baglar.
• GDPbaglamış formu inaktif,GTP bağlamış
formu aktiftir.
• GTPaz aktivitesina sahip moleküllerdir.
• G Proteinlerinin çeşitleri;
Trandusin-C-GMP Fosfodiesteraz
ivasyonu
Gpı- Fosfolipaz C Aktivasyonu
©2002 Lee Bardwell
G proteinlerinin çeşitleri
Gs→ s→AC→cAMP↑
Gi→ i→AC→cAMP↓
Gq→ q →PI-PLC(fosfolipaz )→IP3+DAG
Go→ o→ion channel
Gt→ t →cGMP PDE→cGMP→
Rodopsin
G proteini
Adenil siklaz aktivasyonu
cAMP artışı (ATP den sentez)
Protein kinaz A aktivasyonu
Seçilmiş proteinlerin serin ve treonin
kalıntılarının fosforillenmesi,biyolojik
etkiler
birçok proteinlerin Ser/Thr
rezidülerinin Fosforilasyonu
(1)metabolizmanın
regülasyonu
(2)gen ekspresyonun
regülasyonu
• cAMP protein kinaz A’ya
bağlanır
• Katalitik ünitesi ayrılır
• Nukleusa girer
• cAMP cevap elementi
(CRE) dizisini içeren
genlerin
transkripsiyonunu
aktifler
• CRE bağlayan protein=
CREB’i fosforilleyerek
CRE : cAMP response element
(TGACGTCA)
CREB: CRE binding protein
FOSFOLİPİDLER
• Hücre içi sinyal iletiminde sıklıkla kullanılan
yolaklardan birisi, fosfatidil inozitol 4,5bifosfat (PIP2, bir zar fosfolipidi)’dan köken alan
ikinci mesajcıların kullanımı esasına dayanır.
• PIP2, plazma zarındaki çift tabaka fosfolipidlerin
iç bölümünde yerleşmiştir.
FOSFOLİPİDLER
• Çok farklı hormonlar ve büyüme faktörleri,
fosfolipaz C tarafından PIP2 hidrolizini uyararak,
iki farklı ikincil mesajcı oluşumuna neden olurlar:
• 1- Diaçilgliserol
• 2- İnozitol 1,4,5 trifosfat (IP3)
Fosfolipaz C (β)’nın
protein-tirozin kinazlar ile aktivasyonu:
• PIP2 hidrolizi, G protein-aracılı reseptörlerin ve reseptör
protein-tirozin kinazların aktive edilmesi ile gerçeleşir.
• G protein-aracılı reseptörler ile fosfolipaz C-β (PLC-β)
uyarılır.
PROTEİN KİNAZ C
• PIP2 hidrolizi ile üretilen diaçilgliserol,
protein kinaz C ailesinde yer alan, hücre
büyüme ve farklılaşmasının kontrolünde
önemli roller oynayan, protein-serin/treonin
kinazları aktive eder.
• Örnek: Forbol esterlerinin etkisi.
IP3 ve Ca2+ SALINIMI:
• PIP2 hidrolizi ile oluşan ve sitozole salınan
bir diğer ikinci mesajcı (küçük, polar bir
molekül) IP3, hücre içi depolardan Ca2+
salınım sinyalleri oluşturur.
IP3 ve Ca2+ SALINIMI:
• IP3 ligand-kapılı Ca2+ kanallarına bağlanarak,
endoplazmik retikulumdan Ca2+ salınımını
sağlar.
• Sonuç olarak, sitozoldeki Ca2+ düzeyinin 1 µM’a
(normalde 0.1 µM düzeyinde tutulur) kadar
artması, protein kinaz ve fosfatazları da içeren
çok sayıda farklı hedef proteinin aktivitesini
etkiler.
IP3 ve Ca2+ SALINIMI:
• Birçok hücrede, IP3 üretimi sonucunda hücre-içindeki
Ca2+’un geçici artışı, plazma zarındaki kanallar yolu ile
hücre-dışındaki Ca2+’un içeriye girmesine neden olarak,
daha uzun süreli artışı tetikler.
• Hücre-dışındaki
Ca2+’un
girişi
aynı
zamanda
endoplazmik
retikulumdaki
Ca2+
depolarının
yenilenmesini sağlar.
KALMODULİN
• Ca2+’un birçok etkisi Ca2+-bağlayan bir
protein olan kalmodulin tarafından
düzenlenir.
• Kalmodulin,
sitozoldeki
Ca2+
konsantrasyonu 0.5 µM’a kadar arttığı
zaman aktive olur.
KALMODULİN
• Kalmodulin, 4 adet Ca2+ bağlanma bölgesi olan, davul
tokmağı şeklinde bir proteindir.
• Aktif Ca2+/kalmodulin kompleksi, Ca2+/kalmodulin bağımlı
(CaM) protein kinaz gibi çok sayıda hedef proteine
bağlanır.
2+
Ca /KALMODULİN
BAĞIMLI
PROTEİN KİNAZLAR:
• Aktive olan Ca2+/kalmodulin, protein kinazları da
içeren çok farklı hedef proteine bağlanır:
• Miyozin hafif-zincir kinaz,
• Metabolik enzimler,
• İyon kanalları,
• Transkripsiyon faktörlerinin fosforillenmesi ile
gen ekspresyonunun düzenlenmesi.
cAMP ve
Sinyal İleti
Yolakları Arasındaki İlişki:
2+
Ca
•
Extrasellüler sinyal molekülünün G proteinine bağlı reseptöre bağlanması, adenilil
siklaz aktivasyonuna ve cAMP konsantrasyonunda artışa yol açar.
•
cAMP konsantrasyonundaki artış, sitozolde PKA’yı aktive eder ve katalitik alt
birimlerini serbest bırakır. Daha sonra aktive PKA, CREB gen düzenleyici proteini
fosforile ettiği nukleusa doğru hareket eder.
•
Bu sinyal yolağı, beyinde uzun dönem hafıza için gereksinim duyulan proteinlerin
üretimi için, endokrin hücrelerde hormon sentezini düzenleyen hücrelerdeki birçok
süreci kontrol eder.
IP3 and Protein kinase C have
different effects in different cells
a. IP3
i. Vascular smooth muscle
ii. smooth muscle
iii. skeletal muscle
iv. blood platelet
contraction
contraction
contraction
aggregation of platelets
b. Protein Kinase C. cell growth, differentiation (development into
different tissue types), metabolism.
i. blood platelets
ii. mast cells
iii. smooth muscle
iv. nerve cells
v. adipose tissue
vi. liver cells
serotonin release
histamine release
contractility
neurotransmitter release
fat synthesis
glycogen hydrolysis