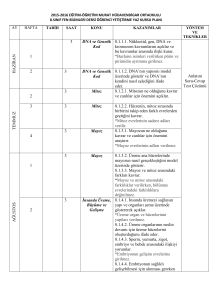
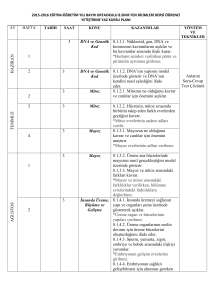
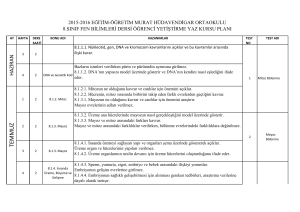
HÜCRE DÖNGÜSÜ
Biyolog Bahattin KARLANKUŞ
CUMHURİYET LİSESİ
Sultangazi
HÜCRE BÖLÜNMESİ
Hücre Bölünmesi
• Tek hücreli ve çok hücreli
organizmaların
– Büyüme
– Gelişme
– Çoğalmaları hücre bölünmesiyle sağlanır.
Hücre Bölünmesi
• Prokaryotik ve ökaryotik hücreler DNA
sentezinin koordinasyonu ve DNA’nın
eşit olarak yavru hücrelere
paylaştırılması yönünden birbirinden
farklılık gösterirler.
• Hücre bölünmesi iki tiptedir.
– Mitoz: diploid somatik hücrelerde genetik
özdeşlik
– Mayoz: haploid germ hücrelerini meydana
getirir
Hücre Bölünmesi
•
Prokaryotlar ve ökaryotlar arasındaki
hücre düzeyindeki farklılıklara rağmen
hücre bölünme süreçlerinde birçok ortak
özellik bulunmaktadır.
–
–
–
–
Hücrenin büyümesi
DNA’nın replikasyonu
Orjinal ve kopyasının ayrılması
Sitoplazmanın bölünmesi
Hücre Bölünmesi
• Bakterilerde hücre büyümesi ve
replikasyon hücre siklusunun büyük bir
bölümünde yer alır ve iki katına çıkan
kromozomlar yavru hücrelere
sitoplazmayla birlikte ayrılır.
Hücre Siklusu
• Hücreler içeriklerini iki katına çıkararak
ve ikiye bölerek çoğalırlar. Bu süreç
HÜCRE SİKLUSU (DÖNGÜSÜ) olarak
adlandırılır.
Prokaryotik Hücre Bölünmesi
•
•
•
•
•
•
Prokaryotlar organizazyon
bakımından ökaryotlara göre daha
basittir.
Ökaryotlar bir çok organel ve fazla
sayıda kromozoma sahiptir.
Prokaryotik hücrelerin bölünmesi
genel olarak “binary fission” ikiye
bölünme şeklinde olmaktadır.
Prokaryotik kromozom tek bir
DNA molekülüdür.
Önce replikasyona uğrar ve daha
sonra kromozomun her bir kopyası
hücre membranının farklı bölgesine
tutunur. Hücre uçlardan çekilmeye
başlayınca orjinal ve kopya
kromozomlar ayrılırlar.
Sitolazmanında ikiye ayrılmasıyla
(sitokinez) genetik içerik
bakımından tamamıyle birbirinin
aynı olan iki yeni hücre meydana
gelir ( düşük oranda da olsa) eğer
kendiliğinden mutasyon olmadıysa)
• Prokaryot kromozomu
araştırmak ökaryotik
kromozoma göre daha kolay
olduğundan prokaryotlarda
genlerin yerleşimi ve
kontrolu hakkında daha
fazla bilgi bulunmaktadır.
• Bu eşeysiz çoğalmanın bir
sonucu, kolonideki tüm
organizmaların genetiğinin
aynı olmasıdır. Bir
bakteriyel hastalığı tedavi
eden bir ilaç kullanıldığında
bu ilaçla karşılaşan koloninin
diğer üyeleride ölecektir.
ÖKARYOTİK HÜCRE BÖLÜNMESİ
• Ökaryotlarda hücre siklusu çok
daha karmaşıktır ve 4 farklı
fazdan oluşur.
– S fazı: DNA sentezi
– G1 (ilk aralık) RNA ve protein
sentezi, DNA için sentez
hazırlığı
– G2 (ikinci aralık) DNA sentezi
tamamlanır, RNA ve protein
sentezi devam eder.
– S+G1+G2 = İNTERFAZ
Hücre Siklusu
• Hücreler interfaz süresince sabit bir hızda büyür.
Bölünen hücreler için iki katına çıkma süresi iki mitoz
arasında geçen süredir.
• Sitokinezle sonlanan mitozdan sonraki faz DNA sentezi
ile mitoz arasındaki G1 fazıdır.(gap1) Bu faz sırasında
hücre metabolik olarak aktiftir ve sürekli olarak büyür
ve DNA sentezi için gerekli proteinleri sentezler, fakat
DNA’sının sentezini yapmaz.
• G1 fazından sonra DNA replikasyonunun yer aldığı S
fazı (sentez fazı) gelir.
• DNA sentezinin tamamlanmasını G2 fazı (gap2) izler.
Bu fazda hücre büyümeye devam eder ve mitoz için
gerekli olan proteinler sentez edilir.
Hücre Siklusu
• Hücre siklusu fazlarının süreleri farklı hücre tipleri
için oldukça çeşitlilik gösterir.
• Toplam hücre siklusu 24 saat olan hızlı çoğalan bir
insan hücresinde
G1 fazı 11 saat,
S fazı 8 saat,
G2 4 saat ve
M fazı 1 saattir.
– Diğer hücreler örneğin maya hücresi
tüm siklusunu yaklaşık 90 dakikada
tamamlayabilir.
Hücre Döngüsü
• Daha da kısa hücre siklusları yumurtanın döllenmesinden
sonraki erken embriyonik hücrelerde 30 dakika olabilir.
Bu tip hücrelerde hücre büyümesi olmaz yumurta
sitoplazması küçük hücrelere hızlı bir şekilde bölünür ve
G1 ve G2 fazları yoktur. DNA sentezi hızlı bir şekilde
çok kısa olan S fazında gerçekleşir
Ökaryotik hücre siklusu
• Hücre büyümesi kesintisiz bir süreç olmakla
beraber DNA, hücre siklusunun sadece bir fazında
sentez edilir ve replikasyona uğrayan kromozomlar
daha sonra bir seri karmaşık işlemi kapsayan hücre
bölünmesi ile yavru nukleuslara bölünür.
• Hızlı çoğalan embriyonik
hücrelerin dışında yetişkin
hayvanlarda bazı hücreler
bölünmeyi tamamıyla durdurur
(sinir hücreleri, kas hücreleri)
ve bazıları da sadece nadiren
hücrenin yaralanması veya
ölümüyle kaybolması sonucu
yerine koymak için gerekli
olduğunda bölünür. Bu ikinci tip
hücreler deri fibroblast
hücreleri ve karaciğer, böbrek
ve akciğer gibi iç organ
hücreleridir. Bu hücreler
metabolik olarak aktif
oldukları, uygun bir hücre dışı
sinyal almadıkça çoğalmadıkları
bir faz olan G0 fazına girmek
üzere G1 fazından ayrılırlar
(çıkarlar).
Hücre Siklusu Kontrolu
• Hücre siklusunun
aşamaları arasındaki
geçişler korunmuş bir
düzenleme
mekanizması
(“checkpoints” sistemi)
tarafından kontrol
edilir. Bu mekanizma
sadece hücre
siklusunun farklı
olaylarını kontrol
etmez aynı zamanda
hücre çoğalmasını
kontrol eden hücre dışı
sinyallerle hücre
siklusu arasındaki
ilişkiyi de sağlar.
Hücre Siklusu Kontrolu
• G1 evresindeki karar verme
noktası (“Restriction point”
sınırlayıcı nokta) hayvan
hücresinin çoğalmasını
düzenler. Mayalara zıt
olarak hayvan hücrelerinin
hücre siklusunu
tamamlamaları öncelikle
besin maddelerinin
kullanılabilmesinden çok
hücre dışı “growth” büyüme
faktörleri tarafından
düzenlenir.
Hücre Büyümesi ve Hücre dışı
Sinyallerle Hücre siklusunun
Düzenlenmesi
• Hücre siklusunun bölümleri arasındaki geçiş ve
farklı hücre sikluslarında meydana gelen
çeşitli işlemler hücre içi sinyallerin yanı sıra
çevreden gelen hücre dışı sinyallerle de
düzenlenir.
• Hücre dışı sinyallerle hücre siklusunun
düzenlenmesine bir örnek hayvan hücrelerinin
çoğalmasına büyüme faktörlerinin etkisidir.
• Hücre büyümesi, DNA replikasyonu ve mitoz
gibi hücresel süreçler hücre siklusu sırasında
farklı kontrol noktalarında düzenlenirler.
Hücre Siklusu
• Büyüme (“Growth”) Faktörleri
– “Platelet-derived Growth factor” (PDGF): Bağ doku ve nöroglia
hücrelerinin çoğalmasını
– “Epidermal Growth Factor” (EGF):Birçok hücre tipinin
çoğalmasını
– “İnsulin like Growth factor” (IGF-1): yağ hücreleri ve bağ doku
hücrelerinin çoğalmasını
– “Trasforming Growth Factor” (TGF): Hücre tipine bağlı olarak
hücrelerin farklılaşmasını düzenler
– “Fibroblast Growth Factor” (FGF): Fibroblast ve endotel
hücrelerin çoğalmasını
– “Nerve Growth FActor” (NGF): Bazı duyu ve merkezi sinir
sistemi nöronlarının hayatta kalma süresini uzatır.
– İnterlökin 2: T lenfositlerim çoğalmasını
– “Hemopoietic Cell Growth Factors” : Kan hücrelerinin oluşumunu
etkiler
• Birçok hücre tipinde hücre siklusu ana kontrol
noktası, G1’den S fazına geçişi kontrol eden geç G1
noktasıdır. Bu düzenlenme noktası ilk kez
Saccharomyces cerevisiae’de saptanmıştır. START
olarak bilinen bu noktayı geçen hücreler S fazına
girerler ve hücre bir hücre bölünmesi geçirir.
•
•
START noktasının geçilmesi oldukça yüksek düzeyde kontrol edilen bir
süreçtir. Besin maddelerinin varlığı, hücrenin boyutu gibi hücre dışı
sinyallerle kontrol edilir. Örneğin eğer maya besin açlğı ile karşılaşırsa
hücre siklusunu START noktasında durdurur ve S fazına geçmeden uyku
(dinlenme) durumuna geçer.
Maya eşleşmesinde etkili olan polipeptit faktörler de hücre siklusunu
START noktasında durdurur. Bu faktörler maya hücresini S fazına
sokmak yerine haploid maya hücrelerinin birbiriyle birleşmesini sağlar
• Hücre dışı sinyallerin izlenmesinde bir karar noktası
gibi iş görmesine ek olarak START noktası hücre
çoğalmasının; DNA replikasyonu ve hücre bölünmesi ile
koordine olduğu bir noktadır.
• Bu düzenlenme özellikle tomurcuklanan mayalar için
önemlidir. Çünkü ana hücre çok büyük, yavru hücreler
çok küçüktür. Bu hücrelerin tekrar bölünmeden sabit
bir boyda olması için küçük yavru hücrelerin hızlı bir
şekilde en az annesi kadar büyümesi gerekmektedir.
• START noktasındaki kontrol ve düzenleme bölünecek
hücrenin belirli bir büyüklüğe ulaşmasını sağlar.
• Bu düzenlenme her bir hücrenin START noktasını geçmeden
önce minimum boyuta ulaşmasını gerektiren kontrol
mekanizmasıyla birlikte olur. Sonuç olarak küçük yavru hücre
G1’de daha uzun zaman harcar ve ana hücreden daha fazla
büyüyerek tekrar bir tomurcuk oluşturma konumuna girer. Bu
nedenle mayada G2 fazı oldukça kısadır.
Hücre siklusu kontrolu
•
•
•
Birçok hücrenin çoğalması G1 fazında
düzenlenmekle beraber bazı hücrelerin
çoğalmasının kontrolu özellikle G2 fazında
yapılır. Buna bir örnek
Schizosaccharomyces pombe (“fission
yeast”) mayasının hücre siklusudur. G2
fazından M fazına geçişte hücre boyutu
ve besin varlığının etkili olduğu nokta
temel kontrol noktasıdır. Bu maya heriki
ucundan uzayarak büyür ve hücrenin
ortasında hücre duvarı oluşturarak ikiye
bölünür. S.cerevisiae’den farklı olarak
normal G1,S,G2 ve M fazlarına sahiptir.
Sitokinez G1 de olur. Hücre boyutu
(uzunluğu) hücrenin siklusun hangi
aşaşmada olduğunu gösterir.
Hayvanlarda hücre siklusunun G2 fazında
kontroluna örnek oositlerdeki kontrol
mekanizmasıdır. Omurgalı oositleri G2
fazında çok uzun yıllar kalabilirler. M
fazına geçişleri hormonol uyarılma ile
birlikte olur.
SONUÇ : Hücre dışı sinyaller hücre
siklusunda G2 fazından M fazına ve G1
den S fazına geçişleri düzenleyerek hücre
çoğalmasını kontrol eder.
Schizosaccharomyces pombe’nin
birbirini izleyen mitoz ve sitokinez
aşamalrının mikroskobik görüntüsü,
Robinow
•
•
•
•
Hücre Siklusu Kontrol Noktaları
Hücre siklusunun farklı fazları
arasındaki koordinasyon, hücre
siklusunda bir fazın işlemi
tamamlanmadan bir sonraki faza
geçişini engelleyen kontrol noktalarına
ve “feedback” kontroluna bağlıdır.
Bazı kontrol noktaları tamamlanmamış
veya zarar görmüş kromozomun
replikasyonunun yapılmamasından ve
yavru hücreye geçmemesinden
sorumludur.
Bu noktalardan en iyi bilineni G2’dedir
ve DNA replikasyonu tamamlanmadan
mitoza girişi engeller. Bu G2 kontrol
noktası replikasyon olmamış DNA’ya
duyarlıdır. Böyle bir DNA hücre
siklusunu durdurmaya yol açan bir sinyal
oluşturur. Böylece G2 kontrol noktası S
fazı tamamlanmadan önce Mitoza girişi
engeller.
G2’deki kontrol noktası zarar görmüş
DNA’ya karşı da duyarlıdır. Bu noktada
hücre siklusu durdurularak zarar
görmüş DNA’nın onarılması için zaman
kazanılır.
• Her bir hücre döngüsü için DNA
replikasyonunu bir kereyle sınırlayan
moleküler mekanizmada, MCM proteinleri
başlangıç
noktasına
replikasyon
kompleksi (ORC) ile birlikte bağlanır.
• MCM
proteinleri,
kopyalamanın
başlamasına izin veren “yetki faktörleri”
olarak davranırlar.
• DNA’ya
bağlanmaları
G1’de
bağlanabilecek şekilde düzenlenmiştir ve
DNA replikasyonunun, sadece hücre S
evresine girdiğinde başlamasına izin verir.
Başlama gerçekleşince MCM proteinleri
orijinden ayrılır, böylce hücre mitozdan
geçip bir sonraki hücre döngüsünün G1
evresine girene kadar kopyalama tekrar
başlatılamaz.
Hücre Siklusu Kontrol Noktaları
• DNA hasarı hücre siklusunu
sadece G2’de tutmaz aynı
zamanda S fazından
hücrenin çıkışını da
yavaşlatır ve G1’deki kontrol
noktasında hücre siklusunun
ilerlemesini durdurur.
• G1 tutuklanması, hasarlı
DNA’nın replikasyonunun
yapılacağı S fazına
girmeden önce hatanın
onarılmasına izin verir.
Hücre Siklusu Kontrol Noktaları
• S’deki kontrol noktası
hasarlı DNA’nın replike
olmadan onarılması için
DNA’nın bütünlüğünü
sürekli izler.
• DNA replikasyonu
sırasında yanlış baz
eşleşmesi ya da DNA
bölümlerinin eksik
replikasyonu gibi
oluşabilecek hataları
bulma ve onarma şeklinde
kalite kontrol görevi yapar
Hücre Siklusu Kontrol Noktaları
• Bir başka hücre siklusu
kontrol noktası
kromozomların mitoz
sırasında dizilimlerini
kontrol eder.
• Böylece tam bir kromozom
takımının yavru hücrelere
geçirilmesi sağlanır.
• Eğer hatalı dizilim olursa
mitoz fazının metafazında
siklus, kromozomların tam
bir takımının yavru
hücrelere dağılımı
düzenleninceye kadar
durdurulur.
Hücre Siklusu Kontrol Noktaları
• G2 kontrol noktası S fazı tamamlanmadan mitoza
girişi engeller. Böylece replikasyonu tamamlanmamış
DNA yeni yavru hücrelere geçmez
• En az bunun kadar önemli olan bir diğer nokta
genomun her bir hücre siklusunda sadece bir kere
replikasyon olmasıdır. Böylece DNA bir kere
replikasyon olduktan sonra mitozdan önce yeni bir S
fazına girişi engelleyen kontrol mekanizmaları
olmalıdır. Bu kontrol, hücrelerin G2’de tekrar S fazına
girmesini engeller ve mitozdan önce bir başka DNA
replikasyon döngüsüne girmesini durdurur.
G1, S ve G2’deki
kontrol
noktalarında
hücre
döngüsünün
durdurulması:
Algılayıcı
proteinler
kompleksi hasarlı ya da
replike olmamış DNA’ya
bağlanarak ATM ve ATR
protein kinazları aktive eder.
ATM ve ATR sırasıyla Chk2
ve Chk1 protein kinazları
fosfatlayarak aktive eder ve
hücre
döngüsünün
durmasını sağlar.
•
•
•
Araştırıcılar (Potu Rao ve Robert Johnson, 1970) siklusun farklı
fazlarındaki hücreleri izole ettikten sonra hibrit hücreler elde etmek
için hücreleri birbirleriyle birleştirdiler.
S fazı hücreleri G1 hücreleri ile birleştirildiğinde G1 nukleusu hemen
replikasyona başlar. S faz hücreleri G2 hücreleri ile birleştirilirse sadece
S faz nukleusu DNA replikasyonuna devam eder.
G2 nukleusunun, M fazını geçmek zorunda olduğu ve birbaşka DNA
replikasyonuna başlamadan önce G1 fazına girmesi gerektiği ortaya
çıkmaktadır.
• SONUÇ: Mitoz oluşuncaya kadar
G2 nukleusunda kontrol
mekanizması DNA
replikasyonunu durdurmaktadır
Hücre Siklusu Kontrol Noktaları
• Memeli hücrelerinin G1
kontrol noktasında
durması p53 proteini
olarak bilinen protein
aracılığı ile olmaktadır.
Bu protein hasarlı DNA
tarafından çok hızlı bir
şekilde indüklenir.
Hücre Siklusu Kontrol
Noktaları
• Bir çok kanser tipinde p53 proteinini şifreleyen gende
mutasyon saptanmıştır.
• Bu mutasyonların sonucu olarak p53 işlevinin kaybolması
DNA hasarına cevap olarak G1 de durma olayı
gerçekleşmez. Böylece hasarlı DNA replikasyona uğrar
ve yeni yavru hücrelere onarılmadan geçer. Hasarlı
DNA’nın bu şekildeki kalıtımı mutasyon sıklığının ve
kanser gelişimine eşlik eden hücre genomunun genel
kararsızlığının artmasına neden olur.
• P53 genindeki mutasyonlar insan kanser tiplerinde en
sık görülen genetik değişikliklerdir.
• SONUÇ: Çok hücreli organizmaların hücre siklusunun
kontrolu ÇOK ÇOK ÖNEMLİDİR !!!!!
Hücre siklusu fazlarının belirlenmesi
• Mitoz fazındaki hücreler mikroskobik olarak kolaylıkla
ayırt edilebilir.
• Siklusun diğer fazları (G1, S ve G2) biyokimyasal
kriterlerle ayrılır. S fazındaki hücreler DNA sentezi
sırasında kullandıkları timidinin radyoaktif işaretli
olmasıyla ayrılırlar. Örneğin hızlı çoğalan bir insan
hücresi populasyonunda kültür kısa bir süre (15
dakika) radyoaktifli timidin etkisinde bırakılıp
otoradyografi ile analiz edilirse hücrelerin yaklaşık
1/3 ünün işaretlendiği görülür ve hücrelerin S fazında
olduğu anlaşılır.
•
Hücre siklusunun farklı
aşamalarındaki hücreler DNA
içeriklerine göre de ayırt
edilebilir. Örneğin hayvan
hücreleri G1 fazında 2n’dir. S
fazının sonunda DNA içeriği 4n
olur. Hücreler S fazında 2n-4n
arasında DNA içeriğine
sahiptir. G2 ve M fazlarında 4n
olarak kalır Sitokinez sonrası
DNA miktarı tekrar 2n olur.
Deneysel olarak DNA
miktarındaki bu değişim DNA’ya
bağlanan özel fluorosan
boyaların “flow cytometer” ile
tek bir hücredeki fluorasan
yoğunluğun ölçülmesiyle
saptanır ve hücreler G1, S ve
G2/M fazlarında ayrılırlar.
Hücre Döngüsü Gelişiminin Düzenleyicileri
• Hücre
döngüsünün
düzenlenmesi ile ilgili
şu anki bilgilerimiz:
maya,
deniz
kestanesi, kurbağa ve
memelilerden
elde
edilmiştir.
• Ökaryotların
hücre
döngülerinin
korunmuş bir seri
protein
kinaz
tarafından
kontrol
edildiği gösterilmiştir.
Hücre Siklusunun Düzenlenmesinde
etkili olan moleküller
• Ökaryotik hücrelerin hücre siklusunun
kontrolünde protein kinazlar görev alır.
• Kontrol noktalarını geçişte önemli rol
oynarlar.
• Mayalarla yapılan çalışmalar sonucunda
işlevleri ortaya çıkarılmıştır.
• MPF: Cdc2 ve Siklin Dimeri
Olgunlaşmayı ilerleten faktör. G2’den M’ye
geçişte genel düzenleyici görevi vardır.
• Siklin ve Siklin Bağımlı Kinaz Aileleri
Cdc2 ve siklin B birbiriyle ilişkili geniş ailelerin
üyeleridir. Bu ailelerin farklı üyeleri hücre
döngüsünün farklı evrelerinde kontrolü
sağlarlar.
Hücre Siklusunun
Düzenlenmesi
• Hücre siklus düzenlenmesinde anahtar rol oynayan molekülün
varlığı çeşitli araştırmalarla saptanmıştır. Bu çalışmalardan ilki
kurbağa yumurtaları ile yapılmıştır.
• G2 evresinde bekletilen yumurtalar progesteron hormon uyarısı
alınca Mayoz evresine geçmektedir. G2 de iken progesteron
uyarısı almış ve Mayoz evresine geçmiş yumurtalardan alınan
sitoplazmalar enjeksiyonla horman etkisinde olmayan hücrelere
aktarıldığında bu yumurtaların da Mayoz fazına geçtikleri
gösterilmiştir.
• Hormon tarafından uyarılmış sitoplazmadaki bir faktörün
yumurtayı G2 den M fazına geçirmekten sorumlu olduğu
saptanmıştır.
• Bu faktör “Maturating Promoting Factor” (MPF) olarak
adlandırılmıştır.
Hücre Siklusunun
Düzenlenmesi
• Diğer bir çalışmada Lee Hartwell ve ark. (1970)
S.cerevisiae’de hücre siklus işlemlerinde kusurlu, ısıya
duyarlı bir mutant tanımlamışlar ve “Cell division
cycle” kelimelerinin kısaltılması olan cdc adını
vermişlerdir.
• Bu mutant hücre siklusunu uygun sıcaklık olmadığında
START noktasında durdurmaktadır.
• Cdc 28 olarak adlandırılan proteinin G1 evresindeki bu
kontrol noktasını geçmede önemli olduğu saptanmıştır.
Hücre Siklusunun
Düzenlenmesi
• Paul Nurse ve ark. S.pombe mayasında benzer
bir çalışma yapmıştır.
• cdc2 mutantı hücre siklusunu G1’den S fazına
ve G2’den M fazına geçişte durdurmaktadır.
• cdc28 ve cdc2’nin birbirinin homoloğu olduğu
saptanmıştır.
• Moleküler klonlama ve dizi analizi çalışmaları
bu genlerin bir kinaz şifrelediğini
göstermiştir.
Hücre Siklusunun
Düzenlenmesi
• Tim Hunt ve ark (1983)
deniz kestanelerinin
embriyolarında
interfazda biraraya
gelip mitoz sonuna
doğru ayrılan 2 protein
varlığını saptamışlardır.
• Bu proteinler Siklin’ler
(siklin A ve siklin B)
olarak dlandırılmıştır.
Hücre Siklusunun Düzenlenmesi
• Tüm bu çalışmaların
sonucunda hücre siklusu
kontrol sistemi için bir
kavram geliştirilmiştir.
• 1988 yılında kurbağa
yumurtasından
saflaştırılmış MPF’nin iki
alt birimden oluştuğu
gösterilmiştir.
– Siklin protein = düzenleyici
alt birim ve işlevi için
Cdc2’nin katalitik
aktivitesine gerek duyar.
– cdc 2 = kinaz
Siklin ve Siklin-Bağımlı Kinaz Aileleri
Mayada: Cdc2, hem START’dan geçişi hem de mitoza girişi kontrol eder.
Yüksek ökaryotlarda: Cdc2 ve Cdc2-bağlantılı kinazlar (siklin-bağımlı
kinaz, Cdk) hücre döngüsünün çeşitli aşamalarını kontrol ederler.
Cdk’ların aktivitesi en az dört moleküler mekanizma ile düzenlenir.
siklin sentezi ve
parçalanmasıyla
1.Siklinlerle birleşme
3.Cdk/siklin
kompleksine
inhibitör proteinlerin
bağlanması (CKI)
4.14 ve 15. tirozin
bakiyesinin inhibitör
fosforilasyonu
Wee 1 protein kinaz tarafından
gerçekleştirilir. Özellikle Cdc2 ve
Cdk2 de oluşur. Geriye Cdc25
protein fosfataz ile döner
2.161. Threonin
bakiyesinin aktive
edici fosforilasyonu
CAK (Cdk-aktifleştiren kinaz)
tarafından gerçekleştirilir. CAK:
Cdk7/siklinH. Bu kompleks TFII H
ile de birleşir. Transkripsiyon, DNA
tamiri ve Hücre döngüsünde yer
alır.
Hücre Siklusunun Düzenlenmesi
• Memeli hücrelerinde siklin B
sentezi hücre siklusunun S
evresinde başlar.
• Sentezlenen siklin B cdc 2
bir kompleks oluşturur.
• Bu kompleks hücre
siklusunun S ve G2
evrelerinde iş görür.
Memeli hücrelerinde , iki Cdk inhibitör ailesi, farklı Cdk/siklin
kompleksinin düzenlenmesinden sorumludur.
Böylece, Cdk aktivitesinin düzenlenmesinde ilave bir kontrol
mekanizması oluşur.
Büyüme Faktörleri ve D-Tipi Siklinler
• Hayvan
hücrelerinin
çoğalması, genellikle
G1
evresindeki
Restriksiyon
(R)
noktasında
çeşitli
hücre-dışı
büyüme
faktörleri
tarafından
düzenlenir.
• Büyüme faktörlerinin
yokluğunda, hücreler
G0’a girer. Tekrar
hücre
döngüsüne
girmeleri,
ancak
büyüme
faktörlerinin
• Siklin D regülasyonundaki bozukluklar kanser hücrelerinin
karakteristik özelliği olan kontrolsüz çoğalmaya neden
olur.
• İnsan kanserlerinin
– hücre döngüsü regülasyonundaki bozukluklardan
– Büyüme faktörü reseptörleri tarafından aktive edilen
hücre-içi
sinyal
yolaklarındaki
anormalliklerden
kaynaklanır.
Lenfoma vb.bir çok kanserde, siklinD1’in sürekli sentezi ya
da Cdk4,6/SiklinD’ye bağlanan Ink4 Cdk inhibitörlerini
inaktive eden mutasyonlar bulunmaktadır.
Siklin D ve biyüme faktör kontrolü ile kanser arasındaki
ilişki, Cdk4/siklinD kompleksinde görev yapan Rp
preoteini aracılığıyla kesin ortaya konmuştur.
Rb proteininin DNA sentezi ve hücre döngüsünün ilerlemesi için gerekli
genlerin anlatılmasını sağlayarak hücre döngüsünde anahtar rol oynadığı
ortaya konmuştur. Rb’nin aktivitesi hücre döngüsü boyunca oluşan Rb
fosforilasyonu tarafından düzenlenir. Rb G1 evresinde E2F’ye bağlanarak
anlatımı E2F tarafından düzenlenen genlerin anlatımını baskılar,
Cdk4,6/siklinD ile fosforlanan Rb E2F’den ayrılır ve ilgili genlerin anlatımı
gerçekleşir.
Hücre Döngüsü İnhibitörleri
Hücre çoğalması sadece büyüme faktörleri
tarafından değil, hücre döngüsünü inhibe
edici uyaranlar tarafından da düzenlenir.
Hücrede bir DNA hasarının oluşması, hücre
içi P53 proteinin seviyesinde artmaya neden
olur. P53 de Cdk inhibitörü olan p21’in
sentezlenmesine yol açar. P21 bazı
Cdk/siklin komplekslerini inhibe ederek
hücre döngüsünün durmasına neden olur.
Aynı
zamanda
doğrudan
DNA
replikasyonunu da inhibe ederler. p21 DNA
pol. alt birimi olan çoğalan hücre nükleer
antijeni
(PCNA)’ne
bağlanarak
DNA
replikasyonunu inhibe eder. Böylece hasarlı
DNA’nın replikasyonu engellenir.
S ve G2 kontrol noktalarında replike olmamış ya da hasara uğramış
DNA’ların varlığında, Chk1 ve Chk2 protein kinazlar tarafından, hücre
döngüsünün engellendiği farklı bir mekanizma daha vardır. Chk1 ve Chk2
bir protein fosfataz olan Cdc25C’i fosfatlayarak inhibe ederler. İnaktif
Cdc25C Cdc/siklinB kompleksini defosforilleyemediğinden Cdc2 aktive
olmaz ve hücre G2’de durur.
• 2001 “Physiology or Medicine”
Nobel ödülü Leland Hartwell,
Tim Hunt ve Paul Nurse adlı
araştırıcılara "key regulators of
the cell cycle" çalışmaları için
verildi.
• Genetik ve biyokimyasal
yöntemleri kullanarak bu
moleküllerin siklin ve CDK
olduklarını tanımladılar.
• Bu temel keşif biyoloji ve tıp
alanında derin bir etkiye sahip
olmuştur.
• CDK ve siklinler DNA sentezini,
kromozom ayrılmasını ve hücre
bölünmesini control ve koordine
eden moleküllerdir.
Leland
Hartwell,
1939.
Fred Hutchinson
Cancer Research
Center, Seattle,
WA, USA.
L. Hartwell ve arkadaşları tomurcuklanan maya
(Saccharomyces cerevisiae) ile çalışmışlar. Hücre
döngüsünde kusur bulunan ısıya duyarlı mutant elde
etmişler ve bu mutantlara “cell division cycle mutant”
(cdc) adını vermişler. Bu mutantlar hücre döngüsünün bir
aşamasında durmaktadır. Örneğin cdc28
hücre
döngüsünü START’da durdurmaktadır. Belli ısı koşulu
sağlanmayınca hücre bölünenmemektedir. İlgili proteinin
G1 evresindeki kritik noktayı geçebilmek için gerekliliğini
ortaya koymuştur.
Paul Nurse,
1949.
Imperial
Cancer
Research
Fund,
Lincoln's Inn
Fields,
London, UK.
P. Nurse ve arkadaşları fizyon mayası (Schizosaccharomyces
pombe) ile çalışmışlar. Hücre döngüsünü G1 ve G2’de durduran
cdc2 mutantları elde etmişler. S. cerevisiae’deki cdc28 ile
S.pombe’deki cdc2’nin homolog olduğunu ortaya koymuşlar. Bu
gen ile yapılan moleküler klonlama ve nükleotid dizi analizleriyle
cdc2’nin bir protein kinaz kodladığını ve aynı zamanda insanda da
benzer fonksiyona sahip akraba bir genin varlığını ortaya
koymuşlar. 1987’de ilgili insan geninden CDK1’i izole edip evrimde
korunduğunu gösterdiler. CDK1 siklin bağımlı kinaz olarak
adlandırılan bir ailenin üyesidir. CDK molekülleri hücre döngüsü
boyunca vardır. Ancak aktiviteleri çeşitli siklinlerle birleşmeleriyle
düzenlenir.
Tim Hunt,
1943.
Imperial
Cancer
Research Fund,
Clare Hall
Laboratories,
South Mimms,
UK.
T. Hunt ve arkadaşları deniz kestanesinden (“sea
urchin,Arbacia”) embriyolarıyla çalıştılar. 1982’de deniz
kestanesi ve embriyolarında bir araya gelip ayrışan 2 protein
tanımlamışlar. Bu proteinler interfazda bir araya toplanmakta
ve sonra mitoz sonuna doğru bir birinden ayrılmaktadır. Hunt
bunlara siklinA ve siklinB adını vermiştir.
CDK moleküllerine bağlanan siklinleri keşfettiler. Siklinler
CDK’nın aktivitesini düzenlerler ve fosforlanacak hedef
proteinleri seçerler.
Hunt mitoz boyunca yıkılan siklinlerin hücre döngüsünün
temel bir kontrol mekanizması olduğunu ortaya çıkardı.
MİTOZ
•
•
Tipik bir insan hücresi 24 saatte bir bölünür.
Hücre siklusu iki temel kısımdan oluşur.
– MİTOZ (nukleus bölünmesi) ve İNTERFAZ
– Mitoz ve sitokinez sadece yaklaşık bir saat sürmekle beraber hücre
siklusunun yaklaşık %95’i iki mitoz arasındaki faz olan interfaz ile
geçer
– İnterfaz sırasında kromozomlar yoğunlaşır ve nukleus içinde dağılır
Moleküler düzeyde interfaz hücrenin, bölünmeye hazırlık olarak hücre
büyümesi ve DNA replikasyonu sırasındaki toplam zamandır.
MİTOZ
• Mitoz bir hücresel kopya etkisi yapmak
üzere orjinal kromozomların replikasyonu
ve bölünmesiyle birbirinin tıpa tıp aynısı iki
yavru hücre oluşturma sürecidir.
• Click here to view an animated GIF of
mitosis from
http://www.biology.uc.edu/vgenetic/mitosis
/mitosis.htm.
Mitoz
•
•
•
•
•
•
Yoğunlaşmış replikasyon olmuş kromozomlar önemli yapılara sahiptir.
Kinetokor : İğ ipliklerinin mikrotubullere bağlandığı çok proteinli bir
komplekstir.
Replikasyonu yapılan kromozomlar histon proteinlerle birleşmiş kromatid
olarak bilinen iki DNA molekülünden meydana gelir.
Sentromer: Her iki kromatininde birleştiği noktadır.
Kinetokorlar sentromerin dış tarafındadır.
Kromozomlar ise yoğunlaşmış kromatindir (DNA+histon proteinleri).
• MİTOZ AYGITI İĞ MEKİĞİ
– Kromozomal iğ iplikleri (kinetokor mikrotubulleri)
• Herbir kinetokora yaklaşık 35-40 mikrotubul bağlanır
• Kinetokorlar her kromozomu kutuplara bakacak şekilde yerleştirirler
• Her kromozomu, mitotik iğciği ikiye bölen ekvator düzlemine yerleştirirler
– Devamlı iğ iplikleri (polar mikrotubuller)
Kutuplardan ekvatora doğru uzanırlar
Mitoz Aygıtı
• Mitoz sırasında replikasyonu
yapılan kromozomlar
sitoplazmanın ortasında
yerleşir ve orjinal DNA’nın
bir kopyası yavru hücrelerin
herbirinde olacak şekilde
ayrılır (örneğin 46 kromozom
ile başlanmışsa yavru
hücrelerin her biri 46
kromozom içermelidir).
• Hücreler bunu başarmak için
kromozomları yavru
hücrelerin içine çeken
mikrotubülleri kullanır.
• Hayvan hücrelerinde iğ
iplikleri mikrotubullerinin
oluşumunda etkili olduğu
düşünülen bir çift sentriol
bulunmaktadır. Bitki ve bazı
ökaryotlarda ise sentriol
bulunmaz. Prokaryotlar iğ
iplikleri ve sentriolleri
olmadığından hücre zarının
bölünme sırasında
kromozomları çekme işini
yaptığı tahmin edilmektedir.
• Sentriol içeren hücreler
aster adı verilen sentriolden
hücre zarına doğru uzanan
çok küçük mikrotubullere
sahiptir. Aster iğ
ipliklererini birarada tutan
bir bağ gibi iş görür.
• Metafaz kromozomlarının iki kutbada eşit
uzaklıkta olması kinetokorlarla sağlanmaktadır.
• Herbir kinetokor üzerindeki çekme gücü, o
kinetokorun bağlı olduğu kutba yaklaştıkça
azalır.
• İki kutuptan birine bağlı olan kromozomun bir
kutba çekilmesi zıt bir gücün ortaya çıktığını
gösterir. Bu güçlerin nasıl oluştuğu ve nereden
kaynaklandığı henüz bilinmemektedir.
• Mitozun fazlarını ayırmak bazen oldukça
güçtür.
• Sürecin DİNAMİK olduğu unutulmamalı !!!!
• PROFAZ mitozun ilk aşamasıdır.
• Kromatin yoğunlaşır, nukleolus kaybolur
• nukleus kılıfı erir, sentrioller (eğer varsa) ayrılır
ve kutuplara göç eder, kinetokorlar ve kinetokor
iplikler oluşur ve iğ iplikleri meydana gelir
• METAFAZ: Kromozomlar (bu aşamada sentromerle birarada
tutulan kromatitlerden ibarettir) iğ ipliklerinin kinetokor ipliklere
bağlandığı iğin ekvatoruna (merkezine), doğru göç eder.
• ANAFAZ: Sentromerlerin ayrılmasıyla başlar ve kromozomların
kardeş kromatitleri iğ ipliğinin zıt kutuplarına çekilmesiyle ayrılma
devam eder
• TELOFAZ: Birbirinden ayrılmış kromatitlerin kutuplara
ulaşmasıyla tamamlanır, nukleus kılıfı yeniden oluşur,
kromozomlar kromatin yapısında şekillenir ve
nukleolus yeniden oluşur. Bir hücrenin olduğu yerde
şimdi tamamıyle aynı genetik bilgiye sahip iki küçük
hücre oluşmuştur.
Sitokinez
•
•
•
•
•
•
•
Bu olay genellikle anafazda başlar.
Hücrenin orta bölgesinde hücre zarı, aktin ve miyozin flamentlerden oluşan
kontraktil halkaların etkisiyle iğcik eksenine dik ve iki yeni nukleus arasında
bir oluk oluşturur.
Nuklear ve sitoplazmik bölünme birbiriyle her zaman bağlantılı
olmayabilir.Ör:Drosphila’da embriyonun erken evresinde sitoplazma
bölünmesi olmaksızın 13 nukleus bölünmesi geçirebilir. Daha sonra
nukleusların etrafında sitoplazma bölünmeleri gerçekleşir.
Ayrışma aktin flamentler ile miyozin II’den meydana gelen bir halkanın
kasılması ile gerçekleşir.
Bu halka hücre zarının sitoplazmik yüzeyine bağlanır.
Anafaz başlangıcında henüz bilinmeyen bir mekanizma tarafından hücre
zarı oluğun içine çekilir.
Oluk hücrenin ortasında birleşene kadar derinleşmeye devam eder ve
sonunda zarlar birbirine kaynaşır.
Bitki hücrelerinde sitokinez
• Yüksek bitkilerin hücreleri hücre çeperi ile kuşatılmış olduğundan
sitokinez farklıdır.
• Zar veziküllerinden yeni bir hücre zarı ve hücre çeperi oluşturarak
bölünürler
• Hücre çeperi yeni iki nukleusun arasındaki bir düzlemde oluşur ve
polar fibril kalıntılarıyla birleşerek fragmoplast yapısını oluşturur.
• İlk hücre plağı fragmoplastın iki yanındaki mikrotubulerle ilişkiye
geçen hücre çeperi öncül molekülleri ile dolu olan veziküllerin
ekvator düzlemine taşınmasıyla olur.
• Hücre plağının şekli ve yeri ileride ana hüçre çeperi ile birleşeceği
yer olan hücre zarının hemen altında yer alan aktin mikrotubul
demetinden oluşan preprofaz bandıdır.
• Hayvan
Bitki
MAYOZ BÖLÜNME
Mayoz Bölünme
• Eşeyli üreme sadece ökaryotik
organizmalarda meydana gelir.
• Eşem hücrelerinin (yumurta ve
spermatazoon) gelişimi sırasında
kromozom sayısının yarıya indirilmesini
sağlayan özel bir hücre bölünmesidir.
• Döllenme (fertilizasyon) sırasında iki eşem
hücresinin birleşmesiyle kromozom sayısı
tekrar iki katına geri döner.
• Haploid ve diploid terimleri hücredeki kromozom
takımlarının sayısını ifade eder. İnsan (eşem
hücreleri hariç) hayvanların çoğu ve pek çok bitki
diploittir.
• Diploid = 2n
Haploid = n
• Poliploidi: 2 kromozom takımından daha fazla
kromozoma sahip organizmalar
• Homolog kromozomlar: Aynı genleri taşıyan
kromozomlardır, farklı ebeveynlerden gelirler
• Allel: Homolog kromozomlar üzerindeki herbir
gen için
olan farklılık
Heterozigot Birey
• Mayoz : Özel bir tip nukleus bölünmesi –herbir homolog
kromozomun bir kopyası (her bir genin ya anneye ya da
babaya ait olan kopyası) yeni gamete (eşem hücresine)
geçer.
• Mayoz bölünme iki önemli sonuca neden olur.
– Kromozom sayısı yarıya iner
– Genler arasında rekombinasyonlar (parça değişimi) olur. Bu da
gen havuzunun zenginleşmesini ve genetik farklılaşmasının
oluşumunu sağlar.
• Mitoz: Hücrenin orjinal poliploidi düzeyini korur – örneğin,
bir diploit hücre (2n) iki diploit (2n+2n) hücre bir haploit
hücre iki haploit hücre (n+n) meydana getirir.
• İnsan vücudunda hücrelerin çoğu mitozla üretilir. Bunlar
somatik hücre hatlarıdır. Gametleri verecek hücreler
germ hücre hatları olarak ifade edilir.
• İnsan vücudunda hücre bölünmesi çoğunlukla mitozdur,
mayoz gonatlarda sınırlıdır.
Yaşam Siklusu
• İnsan yaşam siklusunda
diploit faz baskındır
Haploit faz sadece
gametlerde görülür.
• Vücudumuzdaki
hücrelerin çoğu diploittir
ve diploit germ hücreleri
gametleri meydana
getirmek üzere mayoz
geçirir ve bunu
döllenme izler.
Mayozun Fazları
• Mayoz bölünme 2 ayrı bölünmeden oluşur.
• I. Mayoz : Kromozom takımının yarıya inmesi
• II. Mayoz : I. Mayozu izleyen ekvatoryal bölünme
olarak da adlandırılır. Mitoz bölünmede olduğu
gibi kromozomların kardeş kromatitleri farklı
kutuplara giderek ilk oluşan her bir yavru
hücreden 2 yeni yavru hücre daha meydana
gelir. Sonuçta herbiri haploit sayıda kromozom
taşıyan 4 yavru hücre meydana gelir.
I.Mayoz
• Profaz I:
– Oldukça uzun sürer. 5 alt evreye ayrılır.
– Leptoten, Zigoten, Pakiten, Diploten, Diakinez
• homolog kromozomların eşleşmesidir. (henüz
anlaşılamamış bir mekanizmayla) Replikasyon
olmuş kromozomların birbirine bağlanma
sürecine sinapsis denir. Oluşan kromozom her bir
kromozomdan gelen iki kromatitten oluştuğu için
tetrat olarak adlandırılır. Bu oluşum 4 iplikli kalın
bir yapı şeklindedir. Krosingover bu aşamada
meydana gelebilir. Krosing over sırasında
kromatitler kırılır ve farklı homolog kromozomlar
yeniden birleşirler
•
•
•
•
ABCDEFG
ABCDEFG
abcdefg
abcdefg
•
•
•
•
ABCDEFG
ABcdefg
abCDEFG
abcdefg
Mayoz sırasında krosingover sonrası rekombinanat
kromozomların gametlere dağılımı
Profaz I de meydana gelen olaylar mitoz profazıyla aynıdır; (krosingover ve sinapsis
dışında) kromatinin yoğunlaşarak kromozomları oluşturması, nukleolusun erimesi
ve mekik yapısının oluşumu.
• Metafaz I :Mekiğin ekvatoruna tetratların dizilmesiyle başlar.
İğ iplikleri herbir homolog kromozom çiftinin sentromer
bölgesine bağlanır. Diğer olaylar mitozla aynıdır.
• Anafaz I :Tetratların ayrılmasıyla başlar ve iğ iplikleriyle zıt
kutuplara çekilir. Sentromer bozulmadan kalır.
Metafaz I’de kromatidlerin kinetekorları birbirine
birleşik veya bitişiktir. Bu nedenle, iğin aynı
kutupundan gelen mikrotubuller kardeş
kromatidlerin kinetekoruna tutunurken, zıt
kutuplardan gelen mikrotubuller , eş
kromozomların kinetekorlarına tutunur.
Kiyazmalar anafaz I’de açılır ve eş
kromozomlar iğin zıt kutuplarına hareket
ederler
• Telofaz:Mitoz telofazına benzer sadece replikasyon
olmuş kromozom takımının bir seti hücreye geçer.
Türlere bağlı olarak yeni nukleus kılıfı oluşur ya da
oluşmaz. Bazı hayvan hücreleri bu faz sırasında
sentriol bölünmesine sahip olabilir.
• Profaz II: Nukleus kılıfı erir (eğer telofazda
oluşmuşsa) ve iğ iplikleri yeniden oluşur. Diğer
tüm olaylar mitozun profaz ile aynıdır.
• Gerçekte Mayoz II mitoza çok
benzemektedir.
• Metafaz II: Mitoza benzer şekilde kromozomlar
ekvatör bölgesine sentromerleri birbirine zıt
olacak şekilde sıralanır.
• Anafaz II : Sentromerler ayrılır ve kromatitler
(şimdi kromozomlar ) hücrenin zıt kutuplarına doğru
çekilir.
• Telofaz II: tamamıyla mitoz telofazının aynısıdır.
Sitokinez ile hücreler ayrılır.
• Mayoz sonunda haploit sayıda kromozom
taşıyan 4 hücre oluşur !!!!
MİTOZ ve MAYOZ ARASINDAKİ FARKLAR
Mitoz ploidi düzeyi korurken mayoz yarıya indirir. Mayoz sadece çok hücreli
organizmalrın bir kaç hücresinde oluşurken mitoz daha yaygındır.
• Mitoz
• Mayoz
Mitoz
Mayoz
•
Mitoz sonucu oluşan hücreler birbirinin
tam olarak benzeri iken mayoz bölünme
sonucunda hücreler genetik açıdan
farklıdır.
•
Kromozomlar yavru hücrelere rasgele
dağıldığından ve ayrıca aralarında
krosingover oluştuğundan mayoz bölünme
sonucu oluşan her gamet kendine özgü
genetik yapı taşır.
Mitoz ve mayoz arasındaki fark
Gametogenez
•
•
Germ hattı hücrelerinden (diploit, 2n) gamet oluşum sürecidir.
Spermatogenez: gonat olarak bilinen özelleşmiş organlarda bitkilerde
mitoz yoluyla hayvanlarda mayoz yoluyla sperm oluşum sürecidir.
Bölünmeden sonra hücreler sperm hücrelerini oluşturmak üzere
farklılaşma geçirir.
•
Oogenez: Ovaryum olarak bilinen özelleşmiş organlarda bitkilerde
gametofitte mitoz yoluyla hayvanlarda mayoz yoluyla ovum (yumurta
hücresi) oluşum sürecidir.
•
Spermatogenezde tüm 4 mayotik ürün gamet olarak gelişirken oogenezde
sitoplazmanın çoğu tek büyük bir yumurta hücresi içinde yer alır,
diğerleri gelişmez. Tüm sitoplazma ve organeller yumurta hücresinde
toplanır.
•
İnsanlarda erkekler günde 200.000.000 sperm üretirken dişiler herbir
menstural siklusta genellikle bir yumurta üretir
• Spermatogenez:
ergenlik döneminde başlar yaşam süresince
devam eder ve her gün milyonlarca sperm üretimi yapılır. Sperm oluşunca
olgunlaştıkları ve depolandıkları epididiymise hareket ederler.
Oogenez
•
•
•
Ovaryum folikül hücrelerin dış
tabakasıyla çevrelenmiş
gelişmiş bir yumurtanın
oluşacağı pek çok folikül içerir.
Her bir yumurta primer oosit
(yumurta hücresi) olarak
oogeneze başlar.
Bir dişi, doğumuyla bereber her
biri Profaz I’de olan yaşam boyu
yumurta deposunu taşır.
Ergenlikten itibaren menapoza
kadar toplam 400-500 bin
yumurtadan genellikle bir tanesi
gelişmiş yumurta (sekonder
oosit) olarak her ay serbest
duruma geçer.
Oosit mayoz diploten aşamasında
durdurulur ve bu bekleme süresinde oosit
büyür. Oosit daha sonra, hormonal uyarı ile
uyarılır, mayoza yeniden başlar ve ilk
mayotik bölünmeyi küçük bir kutup cismi
oluşturan asimetrik sitokinezle tamamlar.
Çoğu omurgalı oositler döllenme anına
kadar metafaz II’de tekrar durdurulur.
Metafaz II’deki bir yumurtanın sitoplazması iki hücreli embroyonun
bir hücresine mikroinjeksiyonla aktarıldı. Enjekte edilen hücre
metafazda durdurulurken enjekte edilmeyen hücre bölünmeye
devam etti. Metafaz II’deki yumurtanın sitoplazmasında bulunan
bir faktör (sitositatik faktör) enjekte edildiği embriyo hücresinde
metafaz aşamasındaki duraklmayı uyarıyordu.
Somatik hücre klonlamsı
(nukleus transferi)
• Somatik hücre nukleusunun nukleusu çıkarılmış metafaz II oositine
transferiyle somatik hücre vericisiyle aynı genetiğe sahip yeni
bireylerin geliştirilmesi
• Doku farklılaşması sırasında inaktive olan genlerin nukleusun
yeniden programlanması ile tekrar aktive olması TOTİPOTENSİ
durumu
• Yüksek genetik özelliklere sahip çiftlik hayvanlarının çoğaltılması
• Farmasötik ürünlerin üretimi için transgenik hayvanların üretimi
• Soyu Tükenen hayvanların çoğaltılması
• Gen fonksiyonunun çalışılması, genomun yeniden programlanması
gelişimin düzenlenmesi genetik hastalıkların tedavisi ve gen terapi
konularının çalışılması
• 100 klonlanan emriyodan 0-10 gelişimini tamamlayabilmektedir.
• Çalışmalar çok çeşitli faktörler denenerek optimizasyonu sağlama
yönünde yapılmaktadır.
Hücre siklusunun ve kontrolunun kopyalamadaki önemi
• 1938 de Alman embriyolog Hans Spemann "fantastical
experiment“ olarak adlandırdığı deneyde yumurta
hücresinin nukleusunu çıkardı ve başka bir hücrenin
nukleusunu yerleştirdi. Sonuç başarısızdı
• 1952 de iki Amerika’lı bilim adamı, Robert Briggs ve T. J.
King çok ince bir pipet yardımıyla kurbağa
yumurtasından nukleusu uzaklaştırdılar ve nukleusu
çıkarılmış yetişkin bir kurbağanın vücut hücresine
koydular. Sonuç başarısızdı.
• 1984’de Danimarka’lı embriyolog Steen Willadsen, erken
embriyo hücrelerini kullanarak koyun klonlamayı başardı
ve daha sonra domuz ve maymunlarla da aynı sonuçlar
elde edildi.
Hücre siklusunun ve kontrolunun kopyalamadaki önemi
• Önceleri yapılan çalışmalar sadece hayvan embriyo hücrelerinin tüm
bir organizmayı klonlamada etkili olduğunu gösteriyordu.
Farklılaşmış hücrelerin nukleuslarını kullanarak klonlama yapmanın
mümkün olmadığına karar verilmişti.
• Hayvan hücre siklusu ile ilgili araştırmalar yapan İskoçyalı genetikçi
Keith Campell çalışmalarıyla günümüzde bu sonucun değerini
kaybetmesine neden olmuştur.
• 1990’ların başlarında kanser araştırmalarıyla ilerleyen hücre
siklusunun nasıl kontrol edildiğinin anlaşılması hücrelerin koşullar
uygun olmadıkça bölünmediğini ortaya koymuştur.
• Aynı çamaşır makinesinin sıkma fazına geçmeden önce suyun
tamamen boşaldığını kontrol etmesi gibi hücreler de hücre
bölünmesine geçmeden önce gerekli olan herşeyin kontrol altında
olmasını isteyeceklerdir.
• Campbell aynı nedenle yumurta ve verici nukleusun hücre
sikluslarının aynı evrede olmaları gerektiğini düşündü.
Hücre siklusunun ve kontrolunun kopyalamadaki önemi
• Bu önemli nokta bu olayın anahtar
aşamasıydı.
• 1995 de Campbell ve üreme biyologu Ian
Wilmut aç bırakılmış hücrelerden (böylece
hücreleri G1 kontrol noktasında
durdurarak) ilerlemiş embriyonik
hücrelerden koyunu başarıyla klonladılar.
•
• Tek bir hücreden hızlı bir şekilde bölünerek
embriyonun gelişimi ve farklılaşması sağlanır.
Bu düşünce ile pek çok biyolog tarafından saç,
barsak ve deri hücreleri gibi çok hızlı hücre
siklusuna sahip hücrelerin nukleusunun embriyo
gelişimde kullanılabileceği ortaya atıldı.
• Pek çok kez denenen bu teori başarısızlıkla
sonuçlandı.
• Campell ve Wilmut gelişmiş bir koyunun
meme bezi hücrelerinin nukleusunu
VERİCİ nukleus olarak kullandılar.
• Hücreler hücre kültürü ortamında önemli
besin gereksinimleri bakımından aç
bırakılarak büyümeleri ve dolayısıyla
bölünmeleri durduruldu. Böylece dinlenme
durumuna getirilerek hücre siklusu
bakımından senkronize edildi.
• İskoçyada yaygın olarak bulunan “Blackface” dişi
koyununun yumurta hücresinden nukleus çok ince bir
iğneyle çekildi.
• “Fin Dorset” adı verilen başka bir tip koyunun meme
hücrelerinden aynı yöntemle çekilen nukleus nukleusu
çıkarılmış yumurta hücresi (“enucleated” hücre) içine
yerleştirildi.
• Çok kısa bir elektrik şokuyla sitoplazma ve nukleusun
birleşmesi sağlandı.
• Bu şok aynı zamanda hücreleri aktive etmek için tekrar
bölünmeye başlamaları için de yardımcı olmaktadır.
• Bu yeni hücreler “blackface” dişi koyunun çoğalma
hattını oluşturacaktır.
• Roslin enstitüsünden bilim insanları bu operasyonu 276
kez denediler.
• Normal gelişim süreci olan 148 gün sonra Dolly’nin
doğumu gerçekleşti.
• Dolly normal sağlıklı görünüşlü bir “Fin Dorset “tipi
koyundu. Dolly’nin “blackface” siyah yüzlü olmaması
araştırıcıları doğru klon olup olmadığının Parmakizi
yöntemiyle kontrol edilmesi şeklinde yönlendirdi.
• Dolly’nin taşıyıcı anneyle değil nukleusunun alındığı
annesiyle aynı DNA parmak izine sahip olduğu
saptandı.
• Yerleştirilen 29 embriyodan 13’ü uterusa
tutunabildi.
• Bu nedenle sadece bir embriyo gelişimini
tamamlayabildi. Ve Dolly doğdu.
• 1999 da yayınlanan raporda Dolly’nin hücrelerinin
doğum yaşından daha yaşlı olduğu (verici hücre 6
yaşındaki bireyden alınmıştı) bildirildi.
• Dolly doğumdan 6 sene sonra öldü Gerçek yaşı neydi?
• Koyunların normal yaşam süreleri 12-15 yıldır.
• Çalışmalar Dolly’nin hücrelerindeki telomerlerin aynı
yaştaki normal bir koyunun telomerlerinden daha kısa
olduğunu göstermiştir.
• Telomerler herbir bölünmeden sonra kısalır. Birçok
bölünmeden sonra kromozomlar kararsız uçlara sahip
olur hücre bölünmeyi durdurur ve ölür.
Sonuç olarak;
• Son yıllarda yapılan çalışmalara göre
verici hücrenin siklus fazı G0 veya G1
olmalı
• Verici hücrenin tipi
• Verici hücrenin yaşı önemlidir.
Döllenme, metafaz II’den anafaz II’ye geçişi indükleyerek , oosit mayozunun
tamamlanmasına ve ikinci kutup cisminin çıkışına yol açar. Sperm çekirdeğini
yoğunluğunu kaybeder ve böylece döllenmiş yumurta (zigot) iki haploid nükleusa
(dişi ve erkek pronükleus) sahip olur.
Memelilerde iki pronükleus birbirine doğru hareket ederken DNA’larını kopyalarlar.
Sonra dişi ve erkek kromozomları ortak bir iğ üzerine dizilir ve mitoz başlar. Mitozun
ve sitokinezin taqmamlanmasıyla herbir hücrenin diploid genoma sahip lduğu iki
hücreli embriyo oluşur.
İn vitro fertilizasyon
• In Vitro Fertilizasyon Embryo Transfer
(IVF-ET) ilk kez 1978 de İngiltere’de Dr
Edwards (Embriyolog) ve Dr. Steptoe
(Jinekolog) tarafından yapılmıştır.
• Teknik daha sonra embriyologlar ve
fizikçiler tarafından geliştirilmiştir ve tüm
dünyada 20 000 nin üzerinde tüp bebek
doğmuştur.
İn vitro fertilizasyonun aşamaları
• a) Inducing and monitoring the development of eggs in the ovaries
after the administration of fertility drugs.
b) Collecting eggs from the ovaries.
c) Collecting sperm from the husband.
d) Combining the eggs and sperm together in the laboratory to
facilitate fertilization and growth of early embryos. In appropriate
cases, artificial fertilization using intracytoplasmic sperm injection
(ICSI) is employed.
e) Transferring the embryos into the uterus.
f) Supporting the initial growth of the embryos in the uterus with
hormonal treatment.
g) Freezing extra fertilized embryos for transfer in subsequent cycles
should pregnancy not occur after the initial treatment.