BİY 315
Karbohidratların Metabolizması-I
Yrd. Doç. Dr. Ebru SAATÇİ
2008-2009 Güz Yarı Dönemi
Glukoz
Glukoz, karbohidrat
metabolizmasının temel
maddesidir.
D- ve L- izomerleri optikçe
aktiftir. Birinin çözeltisi
polarize ışığın düzlemini
sağa (+) çevirir, diğerinin
çözeltisi ise aynı derecede
sola (-) çevirir.
Karbohidrat Metabolizmasının Başlıca Metabolik Yolları
Türü ne olursa olsun organizmaya giren her karbohidrat
sonunda glukoza çevrilir. Bu nedenle:
“Karbohidrat metabolizması deyince akla glukoz
metabolizması gelir.”
1) Glikojenez: Glukozdan glikojen sentezi.
2) Glikojenoliz: Glikojenin yıkılması. Bu olayın karaciğerdeki son
ürünü glukoz, kas dokusundaki son ürünü glukoz- -6 fosfattır.
3) Glikoliz (Embden-Meyerhof Yolu): Glukozun pirüvat veya
laktata kadar yıkılması.
4) Pirüvat Metabolizması: Pirüvatın asetil- KoA ya dönüşümü.
5) Trikarboksilik Asit Siklüsü (Krebs Siklüsü) : AsetilKoA içindeki asetil kısmının CO2’e parçalanması ve bu sırada
redükte koenzimlerin oluşumu.
6) Pentoz fosfat yolu: Glukozun bir başka şekilde
oksidasyonu ile NADPH ve pentoz sentez yoludur.
7) Glukoneojenezis: Karbonhidrat olmayan maddelerden
glukoz sentezi.
8) Glukuronik asit yolu: Glukozdan glukuronik asit sentezi.
Karbohidratların Sindirimi ve Emilimi
Diyette bulunan karbohidratlar çoğunlukla polisakkarit ve
disakkarit (nişasta, laktoz ve sukroz), daha az oranda da
monosakkarit (glukoz ve fruktoz) formunda bulunurlar.
Karbohidratların bağırsaktan emilebilmeleri için
monosakkarit haline çevrilmeleri şarttır.
Karbohidratların parçalanma işlemi ağızdan itibaren
başlayabilir. Tükrükte bulunan α-amilaz polisakkarit
parçalayan bir enzim olmasına rağmen, çiğneme süresinin
kısalığı nedeniyle fazla etkin değildir. Midenin asit pH sı bu
enzimin daha fazla çalışmasına olanak vermez.
Pankreastan çok aktif bir başka α-amilaz salgılanır.
Gerek tükrük, gerekse pankreas amilazı α- 1,4 glikozit bağına
sahip polisakkaritleri bir bağ atlayarak parçalar ve aktivitesi
zincir sonuna yahut dallanma noktasına 2 glukoz ünitesi
kalınca durur.
Böylece α- amilaz etkisi sonunda yaklaşık olarak;
% 40 oranında maltoz,
% 30 oranında α- 1,4 ve α- 1,6 bağı içeren α- dekstrin
% 25 oranında α- 1,4 bağı ile bağlanmış 3 glukozlu maltotrioz,
% 5 oranında da 4-9 glukoz içeren polisakkarid molekülleri
meydana gelir.
İnce bağırsak mukozasının fırçamsı kenarı tarafından
salgılanan diğer hidrolitik enzimlerle karbohidrat sindirimi
sürdürülür ve tamamlanır.
Bu enzimlerden α-glukozidaz her defasında bir tane
olmak üzere α1→4 bağı parçalar.
α-dekstrinaz α1→6 bağını koparır ve serbest glukoz
üniteleri meydana gelir.
Yine bağırsak mukozası kaynaklı β-galaktozidaz veya
laktaz laktozu, sükraz ise sükrozu parçalar ve sonunda
glukoz, galaktoz ve fruktozdan ibaret monosakkaridler
meydana gelir.
Monosakkaridlerin İnce Bağırsaktan Emilimleri
Bunun için 3 yol vardır. Bunlar aktif transport,
taşıyıcı moleküllerle yürütülen kolaylaştırılmış diffüzyon
ve basit diffüzyon dur.
Monosakkaritlerin transportunda SGLT1 ve GLUT5 isimli
glukoz taşıyıcıları (glucose transporter) rol oynar.
SGLT1 taşıyıcıları, ince bağırsak hücresinin serozal
tarafına yerleşmiş Na+,K+-ATPaz sistemine (sodyum
pompası) bağımlı olarak çalışır ve monosakkarit, enerji
harcanarak düşük konsantrasyondaki bağırsak lümeninden
alınır ve yüksek konsantrasyonlu ekstraselüler ortama
aktarılır.
Glukoz ve galaktoz emilimi tek yönlü olarak çalışan bu
yolla gerçekleşir (SGLT1).
GLUT5 ise sodyum pompasına bağlı olmaksızın,
konsantrasyon farkına dayalı olarak monosakkaritlerin iki
yönlü kolaylaştırılmış diffüzyonundan sorumludur. Früktoz
ve bir kısım glukoz bu yolla taşınır.
Basit diffüzyonda ise taşıyıcı protein yoktur, olayı
yönlendiren konsantrasyon gradientidir. Glukoz ve fruktoz
emiliminde kullanılır.
Sodyum pompası, ATP’nin parçalanması suretiyle elde
edilen enerjiyi sodyumun aktif olarak hücre dışına
(ekstraselüler kompartman) transferi için kullanır ve
böylece bağırsak hücresi sitoplazmasında Na+
konsantrasyonu düşer.
Olayı dengelemek üzere bağırsak lümeninde bulunan
Na+-taşıyıcı protein aracılığı ile bağırsak hücresine
girerken beraberinde glukozu da taşır (simport olayı).
Glukozun böbrek tubuluslarındaki geri emiliminde de
bağırsaktakine benzer olaylar tekrarlanır.
Glukoz Transporter
GLUT1: RBC, damar endotel hücrelerinde, beyin, böbrek, kolon ve
plasentada yer alan ve bu dokularda glukoz yakalanması, bazal glukoz
transportundan sorumlu olan taşıyıcıdır.
GLUT2: Karaciğer, pankreas, ince bağırsak ve böbrek proksimal tüp
hücrelerinde bulunan ve bu dokularda glukozun hızlı yakalanması ve
salınmasını sağlayan taşıyıcıdır.
GLUT3: Glukoza affinitesi en yüksek olan, temel olarak beyin
nöronlarında ve böbrek ve plasentada bulunan ve glukozun nöronlarda
yakalanması ve transportunu gerçekleştiren taşıyıcıdır.
GLUT4: Yağ, kas hücrelerinde bulunan glukozun insülinle uyarılan
yakalanmasını gerçekleştiren taşıyıcıdır. İnsülinle uyarılabilen tek
GLUT budur.
GLUT5: İnce bağırsakta ve böbrekte yer alır. Aynı zamanda fruktozunda
taşınımını gerçekleştirir.
GLUT6: Nonfonksiyonel
GLUT7: Karaciğerde mikrozomal fraksiyonda yer alır. Endoplazmik
retikulumdan glukozun salınımını gerçekleştirir.
Glukoz ve Hücre
Bağırsaklardan emildikten sonra vena porta aracılığı ile
sistemik dolaşıma katılan glukoz, galaktoz ve fruktoz
hücre düzeyinde tek monosakkarit (glukoz) üzerinden
metabolize edilir.
Fruktozun glukoza dönüşümü bağırsak ve
karaciğerde, galaktozun glukoza dönüşümü ise
yalnızca karaciğerde gerçekleşir.
6 CH OPO 2−
2
3
5
O
H
4
OH
H
OH
3
H
H
2
H
1
OH
OH
glucose-6-phosphate
Glukoz-6-Fosfat, glukoz molekülünü hücre içinde tutan
bir tür kapandır ve glukozun hücre dışına çıkışını
engeller.
Hücre içinde glukozun fosforilasyonu için 2 ayrı
enzimin (Hekzokinaz ve Glukokinaz) bulunuşu
glukozun karaciğer ve diğer organlar tarafından
kullanılış önceliğini belirlemeye yöneliktir. Bu uygulama
her iki enzimin Km değerlerinin bilinmesi ile açıklık
kazanır.
Hekzokinazın glukoz için Km değeri 0.05 mmol/L,
glukokinazın ise 10 mmol/L dir
Bu durum glukozun; kan glukozu yüksek iken
karaciğer, kan glukozu düştükten sonra diğer organlar
tarafından kullanılması anlamına gelir.
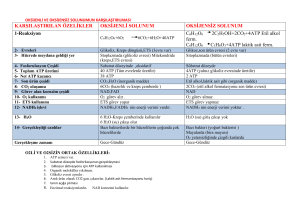
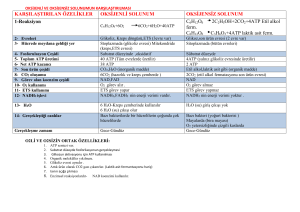
GLİKOLİZ
Glukoz + 2 NAD+ + 2 ADP + 2 Pi Æ 2 pirüvat + 2
NADH + 2 ATP şeklinde reaksiyon özetlenebilir.
Sitoplazmada, sitoplazmik enzimlerle gerçekleşir
Temel amaç enerji elde etmektir.
Aerobik ve anaerobik olarak ikiye ayrılır. Aerobikde pirüvat
TCA siklusuna girer CO2 ve H2O’ya kadar yıkılır. Bu
işlemde NAD ve FAD hidrojenle birleştirilir ve indirgenmiş
NADH+H ve FADH2 elde edilir. Bu moleküller solunum
zincirine H taşırlar. Anaerobik glikolizde ise pirüvat laktata
çevrilir ve son ürün laktattır. Mitokondrisi olmayan
hücrelerde anaerobik glikoliz görülür, çünkü aerobik glikoliz
için gereken enzim sistemleri mitokondride yerleşmiştir.
5
Glikoliz 2 fazdan oluşmaktadır. (a) Hazırlık fazı: Bu fazdan
geçen her glukozdan iki gliseraldehit 3-fosfat oluşur. (b) Hizmet
fazı: Fazın sonundaki ürün pirüvattır.
Hazırlık fazında 2 ATP harcanır; hizmet fazında 4 ATP üretilir.
Net ürün 2 ATP dir.
6 CH2OH
5
H
4
OH
O
H
OH
H
2
3
H
OH
glucose
6 CH OPO 2−
2
3
ATP ADP
H
H
1
OH
5
4
Mg2+
O
H
OH
OH
3
H
2
H
1
OH
Hexokinase H
OH
glucose-6-phosphate
1. Hekzokinaz aşağıdaki reaksiyonu katalizler.
Glukoz + ATP Æ Glukoz-6-P + ADP
Reaksiyon, glukozun 6. C hidroksil O’nin ATP’nin terminal P’na
nükleofilik atak yapmasını içerir.
2+ ile kompleks oluşturarak bağlanır. Mg2+ negatif
ATP, enzime Mg
yüklü fosfat esteri ile etkileşir bu şekilde ATP’nin hekzokinaz enziminin
aktif merkezi için uygun yük konformasyonunu sağlar.
6 CH2OH
5
H
4
OH
O
H
OH
H
2
3
H
OH
glucose
6 CH OPO 2−
2
3
ATP ADP
H
H
1
OH
5
4
Mg2+
H
OH
OH
Hexokinase
O
3
H
H
1
H
2
OH
OH
glucose-6-phosphate
Hekzokinaz’la katalize edilen reaksiyon yüksek derecede
spontandır.
ATP’nin fosfoanhidrit (~P) bağı kırılır.
Glukoz-6-fosfatta oluşan fosfat ester bağı düşük ∆G’ye
sahiptir
6 CH OPO 2−
2
3
5
O
H
4
OH
H
OH
3
H
H
2
H
1
6 CH OPO 2−
2
3
1 CH 2OH
O
5
OH
OH
H
H
4
OH
HO
2
3 OH
H
Phosphoglucose Isomerase
glucose-6-phosphate
fructose-6-phosphate
2. Fosfoglukoizomeraz reaksiyonu:
Glukoz-6-P (aldoz) ÅÆ Fruktoz-6-P (ketoz)
Mekanizma, asit/baz katalizini içerir. Halka açılması enediolat
arametaboliti ile izomerizasyon, ve halka kapanması gözlenir. Enzim
Mg+2 ’a gereksinir.
Phosphofructokinase
6 CH OPO 2−
2
3
O
5
H
H
4
OH
6 CH OPO 2−
2
3
1CH2OH
O
ATP ADP
HO
2
3 OH
5
Mg2+
H
fructose-6-phosphate
1CH2OPO32−
H
H
4
OH
HO
2
3 OH
H
fructose-1,6-bisphosphate
3. Fosfofruktokinaz-1 reaksiyonu:
fruktoz-6-P + ATP Æ fruktoz-1,6-biP + ADP
Hekzokinaz reaksiyonuna benzer bir mekanizmayla reaksiyonu katalizler ve
yüksek derecede spontan bir reaksiyondur.
Glikolizin hız-kısıtlayıcı basamağıdır.
Aktivitesi hücrede ADP ve özellikle AMP fazla miktarda olduğu zaman
artar. Enzim aktivitesi, fruktoz 2,6-biP ve ATP tarafından inhibe edilir
1CH2OPO3
2C
O
HO 3C
H 4C
H
H
2−
H
Aldolase
2−
CH
OPO
2
3
3
+
OH
2C
OH
1CH2OH
2−
OPO
CH
2
3
6
dihydroxyacetone
phosphate
5
C
fructose-1,6bisphosphate
O
O
1C
H 2C OH
2−
3 CH2OPO3
glyceraldehyde-3phosphate
Triosephosphate Isomerase
4. Aldolaz reaksiyonu:
fruktoz-1,6-biPÅÆ dihidroksiaseton-P + gliseraldehit-3-P
Reaksiyon bir aldol kırılımı tepkimesidir (aldol kondenzasyonunun
tersinir tepkimesi)
C atomları aldolaz ürünlerinde tekrar numaralandırılır.
lysine
2−
CH
OPO
2
3
1
H
+
H 3N
C
CH2
CH2
CH2
CH2
+
NH3
COO
2C
−
HO
H
H
3
Enzyme
CH
C
OH
C
OH
4
5
NH (CH2)4
+
2−
OPO
CH
2
3
6
Schiff base intermediate of
Aldolase reaction
Enzimin aktif bölgesindeki Lizin kalıntısı, aktivitede rol oynar.
Fruktoz- 1,6- bifosfat’ın keto grubu, bu lizin kalıntısının e- m
a ino
grubu ile bağlanır ve protone bir Schiff baz ara metaboliti oluşturur.
Bunu takiben C3 ve C4 arasındaki bağ kırılır.
1CH2OPO3
2C
O
HO 3C
H 4C
H
H
5
C
2−
H
Aldolase
2−
CH
OPO
2
3
3
OH
2C
OH
1CH2OH
2−
OPO
CH
2
3
6
fructose-1,6bisphosphate
O
+
O
1C
H 2C OH
2−
OPO
CH
3
2
3
dihydroxyacetone glyceraldehyde-3phosphate
phosphate
Triosephosphate Isomerase
5. Trioz Fosfat İzomeraz (TIM) reaksiyonu:
dihidroksiaseton-P ÅÆ gliseraldehit-3-P
Glikoliz, gliseraldehit- -3P üzerinden devam eder. Bu tepkime ile
glikolizin hazırlık fazı tamamlanır.
Triosephosphate Isomerase
H
H
C
OH
C
O
+
H H
CH2OPO32−
dihydroxyacetone
phosphate
+
H
OH
H H
C
C
+
OH
CH2OPO32−
enediol
intermediate
+
H
O
C
H
C
OH
CH2OPO32−
glyceraldehyde3-phosphate
Bu ketoz/aldoz dönüşümü, bir enediol arametaboliti
üzerinden gerçekleşir. Enzimin aktif bölgesindeki Glu ve His
kalıntıları, kataliz sırasında protonları yapılarına alır ve
bırakırlar.
2-fosfoglukonat enzimin metabolik inhibitörüdür.
Glyceraldehyde-3-phosphate
Dehydrogenase
H
O
NAD+
1C
H
2
C
OH
+ Pi
2−
CH
OPO
2
3
3
glyceraldehyde3-phosphate
OPO32−
+ H+ O
NADH
1C
H
C
2
OH
2−
CH
OPO
2
3
3
1,3-bisphosphoglycerate
6. Gliseraldehit-3-fosfat Dehidrojenaz reaksiyonu:
gliseraldehit-3-P + NAD+ + Pi ÅÆ 1,3-bifosfogliserat + NADH
+ H+
Bu reaksiyon, glikolizin tek NADH açığa çıkan basamağıdır.
H
H
H3N+
C
COO−
CH2
SH
cysteine
O
1C
H 2 C OH
2−
3 CH2OPO3
glyceraldehyde-3phosphate
Enzimin aktif bölgesindeki Cys kalıntısı katalizde
rol alır.
Gliseraldehit-3-fosfatın aldehiti, sisteinin tiyol
grubu ile bir ara form oluşturmak üzere bağlanır.
Aldehit
karbonunun,
karboksilik aside
oksidasyonu
(~ tiyoester), NAD+
yı NADH’a
redükler.
Yüksek enerjili
açil tiyoester’e Pi’ın
bağlanması ile açil
fosfat (~P) ürün
oluşur.
Enz-Cys
Enz-Cys
O
OH
HC
CH
SH
S
OH
OH
CH
CH
CH2OPO32−
glyceraldehyde-3phosphate
CH2OPO32−
thiohemiacetal
intermediate
NAD+
NADH
Enz-Cys
S
O
OH
C
CH
CH2OPO32−
acyl-thioester
intermediate
Pi
Enz-Cys
SH
2−
O3PO
O
OH
C
CH
CH2OPO32−
1,3-bisphosphoglycerate
H
O
H
H
C
C
NH2
+
N
O
−
NH2
+
2e + H
N
R
R
NAD+
NADH
NAD+ redükte olurken, 2 e- + bir H+ (a hidrit)
yapısına alır.
Phosphoglycerate Kinase
O−
OPO32− ADP ATP O
O
C
1C
H 2C OH
2−
OPO
CH
2
3
3
1,3-bisphosphoglycerate
1
Mg2+
H 2C OH
2−
OPO
CH
2
3
3
3-phosphoglycerate
7. Fosfogliserat Kinaz reaksiyonu:
1,3-bifosfogliserat + ADP ÅÆ 3-fosfogliserat + ATP
Reaksiyon sırasında, bir ~P bağı kırılıp tekrar
sentezlendiği için bu fosfat transferi geri dönüşümlüdür.
Enzim, Hekzokinaz gibi substratı tarafından indüklenen
konformasyonel bir değişiklikle aktive olur.
Phosphoglycerate Mutase
O−
O
O−
O
C
C
1
1
H 2C OH
2−
3 CH2OPO3
H 2C OPO32−
3 CH2OH
3-phosphoglycerate
2-phosphoglycerate
8. Fosfogliserat Mutaz reaksiyonu:
3-fosfogliserat ÅÆ 2-fosfogliserat
Fosfat grubu, C2 deki –OH üzerinden, C3 deki –OH üzerine
geçer.
Enzimin aktif bölgesinde bulunan histidin kalıntısının yan
zinciri, fosfatın transferinde donör ve akseptör olarak görev alır.
Reaksiyonun ara metaboliti, 2,3-bifosfogliserat’dır.
histidine
H
H3N+
COO−
C
CH2
C
HN
HC
CH
NH
+
O−
O
C
1
H 2C OPO32−
2−
3 CH2OPO3
2,3-bisphosphoglycerate
Enolase
O−
O
C
C
1
1
H
C
2
O−
O
OPO32−
3 CH2OH
C
2
OPO32− + H2O
3 CH2
2-phosphoglycerate phosphoenolpyruvate
9. Enolaz reaksiyonu:
2-fosfogliserat ÅÆ fosfoenolpirüvat + H2O
Mg++- bağımlı dehidrasyon reaksiyonudur ve fluorit tarafından inhibe
edilir. Fluorofosfat, enzimin aktif bölgesindeki Mg++ ile kompleks
yaparak aktiviteyi inhibe eder.
Pyruvate Kinase
O−
O
ADP ATP
C
1
C
2
O−
O
OPO32−
3 CH2
phosphoenolpyruvate
C
C
1
C
2
O−
O
1
OH
3 CH2
enolpyruvate
C
2
O
3 CH3
pyruvate
10. Piruvat Kinaz reaksiyonu:
fosfoenolpiruvat + ADP Æ piruvat + ATP
Reaksiyon spontan bir reaksiyondur.
Fosfoenolpirüvat (PEP)’ın hidrolizinin ∆G’si, ATP nin
hidrolizininkinden daha yüksektir.
PEP’dan Pi’ın uzaklaştırılması unstabil bir enol oluşumuna yol açar.
Bu ara ürün kendiliğinden keto formu olan pirüvata dönüşür.
Glikoliz Denge Çizelgesi
4 ATP üretilir.
Net ürün glukoz başına 2 ~P dan 2 ATP dir.
Glikoliz - total metabolik yol (H+hariç):
glukoz + 2 NAD+ + 2 ADP + 2 Pi Æ
2 piruvat + 2 NADH + 2 ATP
Anaerobik Glikoliz
Lactate Dehydrogenase
O−
O
C
C
NADH + H+ NAD+
O
O−
O
C
HC
OH
CH3
CH3
pyruvate
lactate
İskelet kasları egsersiz sırasında anaerobik koşullarda çalışırlar, çünkü
aerobik metabolizma ihtiyaç olan enerji miktarını ağır egzersiz sırasında
karşılayamaz.
Anaerobik glikolizde piruvat, devam eden glikoliz için gereken NAD+
nın ortama geri kazandırılabilmesi için laktat’a dönüşür.
Anaerobik koşullarda, Glikoliz ana enerji kaynağıdır.
- 2 mol ATP → Aktivasyon basamakları
+ 2 mol ATP → Fosfogliserat kinaz
+ 2 mol ATP → Pirüvat kinaz
+ 2 mol NADH → Gliseraldehit 3-fosfat dehidrogenaz
- 2 mol NADH → Laktat dehidrogenaz (Pirüvat → laktat)
ANAEROBİK GLİKOLİZ
- 2 mol ATP → Aktivasyon basamakları
+ 2 mol ATP → Fosfogliserat kinaz
+ 2 mol ATP → Pirüvat kinaz
+ 2 mol NADH → Gliseraldehit 3-fosfat dehidrogenaz.
AEROBİK GLİKOLİZ
Fermentasyon
Glyceraldehyde-3-phosphate
Dehydrogenase
H
O
1C
H
2
C
OH
OPO32−
+ H+ O
NAD+ NADH
1C
+ Pi
H C OH
2−
CH
OPO
2
3
3
glyceraldehyde3-phosphate
2
2−
OPO
CH
2
3
3
1,3-bisphosphoglycerate
Anaerob organizmalar, NADH’ı reokisde edecek olan solunum
zincirine sahip değillerdir. NADH’ı başka bir reaksiyon ile okside
etmek zorundadırlar.
NAD+, glikolizde Gliseraldehit- -3 PDehidrojenaz için gereklidir.
Fermentasyon
Pyruvate
Decarboxylase
Alcohol
Dehydrogenase
CO2
NADH + H+ NAD+
O−
O
C
C
O
CH3
pyruvate
H
O
C
CH3
acetaldehyde
H
H
C
OH
CH3
ethanol
Fermentasyonda pirüvat, etanole çevrilir. Açığa çıkan etanol
organizma tarafından dışarı atılır.
Alkol Dehidrojenaz reaksiyonu sonucunda glikolizin sürekliliği
için gerekli olan NAD+rejenere edilir.
Glikoliz, H+hariç:
glukoz + 2 NAD+ + 2 ADP + 2 Pi Æ
2 piruvat + 2 NADH + 2 ATP
Fermentasyon, glukozdan laktata:
glukoz + 2 ADP + 2 Pi Æ 2 laktat + 2 ATP
Anaeroplar fermantasyon ürünlerini (laktat veya etanol) dışarı
atarlar. Glukoz metabolizması sonucu sadece 2 ATP açığa
çıkar.
Aerobik organizmalarda, piruvat, CO2’e kadar okside olacağı
Krebs döngüsüne ve oksidatif fosforilasyona kadar devam
ederek ek ATP oluşumuna katılır.
∆Go'
kJ/mol
∆G
kJ/mol
-20.9
-27.2
+2.2
-1.4
Fosfofruktokinaz
-17.2
-25.9
Aldolaz
+22.8
-5.9
+7.9
negativ
e
-16.7
-1.1
Fosfogliserat Mutaz
+4.7
-0.6
Enolaz
-3.2
-2.4
-23.0
-13.9
Glikoliz Enzim/Reaksiyon
Heksokinaz
Fosfoglukozizomeraz
Triozfosfat İzomeraz
Gliseraldehit-3-P Dehidrojenaz
& Fosfogliserat Kinaz
Piruvat Kinaz
*Values in this table from D. Voet & J. G. Voet (2004) Biochemistry, 3rd Edition, John
Wiley & Sons, New York, p. 613.
Glikolizin Kontrolü
Glikolizdeki spontan oluşan reaksiyonlar:
Heksokinaz, Fosfofruktokinaz & Piruvat Kinaz
Bu enzimlerin katalizlediği reaksiyonlar Glikoliz’in
kontrol basmaklarıdır.
Lokal kontrol, hücre içi substrat veya ara ürün
konsantrasyonlarının glikoliz kontrol basamaklarına etkisini
içermektedir.
Hormonal kontrol, hormonal aktivasyonlar sonucu açığa
çıkan ikincil mesajcıların hücresel reaksiyonları kontrolüyle
olmaktadır.
1.
6 CH2OH
5
H
4
OH
O
H
OH
H
2
3
H
OH
glucose
6 CH OPO 2−
2
3
ATP ADP
H
H
1
OH
5
4
Mg2+
OH
Hexokinase
O
H
OH
3
H
H
2
H
1
OH
OH
glucose-6-phosphate
Heksokinaz’ı kendi ürünü olan glukoz-6-fosfat allosterik
interaksiyonla, unkompetetif olarak inhibe eder.
Glukokinaz, Heksokinazın karaciğerdeki bir diğer formudur. Glukoz için
Km’i yüksek olduğundan, sadece yüksek [glukoz] aktiftir.
Glukokinaz, glukoz-6-fosfat tarafından inhibe edilemez.
Karaciğer Glukokinazı, glukokinaz regulasyon proteini (GKRP)
tarafından inhibe edilir.
Glukokinazın glukoza karşı olan yüksek Km değeri, karaciğerin kan [glukoz]
yüksek olduğu zamanlarda glukozu kandan alarak, glikojen şeklinde depo
etmesine olanak sağlar.
Glycogen
Glucose-1-P
Glucose
Hexokinase or Glucokinase
Glucose-6-Pase
Glucose-6-P
Glucose + Pi
Glycolysis
Pathway
Pyruvate
Glucose metabolism in liver.
2.
Glycogen
Glucose-1-P
Glucose
Hexokinase or Glucokinase
Glucose-6-Pase
Glucose-6-P
Glucose + Pi
Glycolysis
Pathway
Pyruvate
Glucose metabolism in liver.
Glukoz-6-fosfataz, Pi’ın glukoz- -6P’dan salınımı katalizler. Bu şekilde
glukoz karaciğerden salınarak kana karışımasını sağlayarak kan
[glukoz]’nun düzenlenmesinde rol oynar.
Glukokinaz & Glukoz- -6 fosfataz enzimlerinin her ikiside
karaciğerde bulunur, fakat diğer hücrelerde bulunmaz. Bu durum
kan [glukoz]nun sadece KC tarafından düzenlenmesini sağlar.
3.
Phosphofructokinase
6 CH OPO 2−
2
3
O
5
H
H
4
OH
6 CH OPO 2−
2
3
1CH2OH
O
ATP ADP
HO
2
3 OH
H
fructose-6-phosphate
5
Mg2+
1CH2OPO32−
H
H
4
OH
HO
2
3 OH
H
fructose-1,6-bisphosphate
Fosfofruktokinaz (PFK-1), glikolizin hız-kısıtlayıcı basamağıdır of
the Allosterik olarak ATP tarafından inhibe edilir.
Düşük ATP konsantrasyonlarında, ATP enzimin sadece aktif
bölgesine bağlanır.
Yüksek konsantrasyonlarda, ATP aktif bölge dışında düşük- affiniteli
regülatör bir bölgeye de bağlanır. Bu da enzimi, inaktif (sıkı) olan bir
konformasyonel değişikliğe zorlar.
60
low [ATP]
PFK Activity
50
40
30
high [ATP]
20
10
0
0
0.5
1
1.5
[Fructose-6-phosphate] mM
2
PFK
- 1’nın sıkı konformasyonel değişiklik formu, yüksek [ATP] da,
diğer substratı fruktoz- -6P karşı olan affinitesini de azaltır (aktivite
eğrisi sigmoidal eğri şeklini alıyor).
AMP konsantrasyonu, ATP nin hücre içi hidrolizi arttıkça
artmaktadır ve ATP nin PFK üzerine olan etkisini antagonize eder
(azaltır).
Glycogen
Glucose-1-P
Glucose
Hexokinase or Glucokinase
Glucose-6-Pase
Glucose-6-P
Glucose + Pi
Glycolysis
Pathway
Pyruvate
Glucose metabolism in liver.
Glikolizin, [ATP] yüksek olduğu durumlarda PFK-1 ile
durdurulması, glukozun parçalanmasını da durdurur.
ATPnin hücre içi konsantrasyonunun çok yüksek olduğu
koşullarda glukozun parçalanmak yerine, glikojen olarak
depolanması daha yararlıdır.
4. Pirüvat Kinazın Düzenlenmesi
[ATP] = Pirüvat kinazın, substratı olan
fosfoenolpiruvata olan ilgisini azaltarak, allosterik
olarak inhibe eder.
[ATP] = Fosfoenolpiruvata olan ilgisi artar.
Reaksiyon hızlanır.
Asetil-KoA ve uzun zincirli yağ asitleriyle de inhibe
edilir.
Pirüvat Metabolizması
Mitokondriye sahip ve aerobik olan hücrelerde pirüvat
mitokondri içine girer. Bu olay özel bir transport sistemi
ile (kolaylaştırılmış diffüzyon) gerçekleşir. Pirüvat
mitokondride pirüvat dehidrogenaz enzim kompleksi
tarafından oksidatif dekarboksilasyon işlemine tabi
tutularak asetil-KoA ya çevrilir.
Reaksiyon geri dönüşümsüzdür ve oldukça kompleks bir
reaksiyondur. Olayı yürüten pirüvat dehidrogenaz enzim
kompleksi 3 farklı enzimden oluşmuştur. Bunlar:
E1 Pirüvat Dehidrogenaz
E2 Dihidrolipoil transasetilaz
E3 Dihidrolipoil dehidrogenaz enzimleridir.
Olayda görev alan 5 koenzim de sırasıyla şunlardır:
1) Tiyamin pirofosfat (TPPH)
2) KoA-SH
3) Lipoik asit (okside ve redükte formlar)
4) FAD/FADH2
5) NAD+/NADH+H+
TPPH a bağlanan pirüvat,
pirüvat dehidrogenaz
tarafından dekarboksile edilir.
Ürün olarak hidroksietil tiyamin
difosfat meydana gelir. Daha
sonra bu bileşik, bünyesindeki
asetil grubunu dihidrolipoil
transasetilaz yardımıyla KoASH ya aktarır. Bu aktarma
reaksiyonunda okside lipoik asit
koenzim görevi yapar. Ancak
görevi bittiği zaman lipoik asit
redükte forma geçmiştir. Bundan
sonraki reaksiyonlarda ise lipoik
asit tekrar okside forma rejenere
edilirken hidrojen ve
elektronlar sırasıyla FAD ve
NAD+ ye aktarılır. Bu son
aşamadaki işlemler
dihidrolipoil dehidrogenaz
tarafından katalizlenir.
Sitrik Asit Döngüsü (TCA Döngüsü)
Aerobik organizmalarda
enerji veriminin en
yüksek olduğu metabolik
yol sitrik asit
döngüsüdür. Bu
metabolik yol, 3
karboksilli asitler içerdiği
için Tri Carboxylic Acid
siklüsü, kısaca TCA
siklüsü veya siklüs
reaksiyonlarını keşfeden
Sir Hans Krebs in
ismine izafeten Krebs
Siklüsü isimlerini alır.
Döngü reaksiyonları okzaloasetatla başlar ve bir çok ara
üründen sonra yine okzaloasetatla sonuçlanır.
Her dönüşünde 1 asetil-KoA’yı parçalar. KoA-SH ilk
reaksiyonda ayrıldıktan sonra geriye kalan asetil ünitesi CO2 ve
H2O ya ayrışır.
Sitrik asit döngüsü kaynak olarak karbohidrat ve yağlardan
gelen asetil-KoA’lar dışında, amino asitlerden gelen karbon
iskeletleri ile de beslenir.
Tüm reaksiyonları mitokondri matriksinde geçer. Bu
reaksiyonların tüm amacı redükte koenzimler (NADH+H+ ve
FADH2) oluşturmaktır. Bu koenzimler daha sonra ATP
sentezi için mitokondri iç zarındaki elektron transport
zincirine aktarılır.
Döngüde birbirini izleyen 8 reaksiyon vardır.
1. Reaksiyon
Sitrat sentaz tarafından katalizlenen ilk reaksiyonda
okzaloasetat asetil- KoA ile birleşerek sitrat ve KoA-SH’yı
meydana gelir. Bu reaksiyon, kuvvetle ekzergoniktir ve
döngünün kontrolünde önemli bir noktadır.
2. Reaksiyon
Sitrat, akonitaz tarafından
önce cis- akonitata daha sonra
izositrata çevrilir. Ara ürün
olan cis- akonitat reaksiyon
sırasında enzimin aktif
merkezini terketmez, yani
serbestleşmez. Olayın bir
diğer yönü ilk aşamada
reaksiyondan ayrılan suyun
ikinci aşamada tekrar devreye
girmesidir.
3. Reaksiyon
İzositrat dehidrogenaz,
izositratı önce okside eder ve
okzalosüksinat meydana gelir.
Daha sonra aynı enzim
dekarboksilasyon işlemini
gerçekleştirir ve sonuçta α-keto
glutarat oluşur. İzositrat
dehidrogenazın NAD+ ve
NADP+ yi koenzim olarak
kullanan iki izoenzimi vardır.
Birinci izoenzim sadece
mitokonride, ikinci izoenzim
ise hem mitokondri hem de
sitoplazmada bulunur. İzositrat
dehidrogenaz reaksiyonu
irreversibl olup ilk redükte
koenzimlerin oluştuğu evredir.
4. Reaksiyon
α-ketoglutarat
dehidrogenaz
enzim kompleksi αketoglutaratı süksinil- K
oA ya çevirir. Bu reaksiyon tümüyle
pirüvat dehidrogenaz enzim kompleksinin katalizlediği
pirüvatın oksidatif dekarboksilasyonu reaksiyonuna benzer.
Alfa- ketoglutaratın oksidasyon enerjisi süksinil- KoA daki
tiyoester bağında saklı tutulur.
5. Reaksiyon
Süksinat tiyokinaz veya
süksinil-KoA sentetaz ile
süksinil- KoA süksinata
çevrilir. Süksinil- KoA daki
yüksek enerjili tiyoester bağı
enerjisi GDP den GTP
sentezinde kullanılır. Bu olay
substrat düzeyinde bir
fosforilasyondur. Ürün
olarak oluşan GTP,
nükleosid difosfat kinaz ile
ATP ye çevrilir. Net ürünler
süksinat ve ATP dir.
6. Reaksiyon
Süksinat
dehidrogenaz
süksinatı fumarata çevrilir.
TCA
döngüsünün
diğer
enzimlerinin aksine bu enzim
mitokondri
iç
zarına
yerleşmiştir ve koenzim olarak
FAD kullanır. Bu reaksiyonda
oluşan FADH2 hidrojen ve
elektronlarını Koenzim Q
(ubikinon) ya aktardığı için
oluşan ATP her hidrojen çifti
başına 3 değil 2 mol dür.
Aşağıdaki şekilde de görüldüğü
gibi yapı bakımından süksinata
çok
benzeyen
malonat,
süksinat
dehidrogenazın
kompetitif inhibitörüdür.
7. Reaksiyon
Bu aşamada fumarat, fumaraz tarafından L
- malat’a çevrilir.
Stereospesifik olan fumaraz reaksiyonun tersine işlemesi
halinde L
- malatı substrat olarak kullanır.
8. Reaksiyon
TCA siklüsünün son aşamasında -L malat, malat dehidrogenaz
tarafından okzaloasetata çevrilir. Standart şartlarda ileri derecede
endergonik olan bu reaksiyon, okzaloasetatın süratle kullanılarak
konsantrasyonunun düşürülmesi nedeniyle sağa doğru da kolayca
işler.
Pirüvat Dekarboksilasyonunun Kontrolü
Pirüvatın mitokondriye girdikten sonra CO2 ve H2O ya kadar
yıkılması ATP oluşumu ile beraber gider. ATP ve ATP oluşumuna
yol açacak olan her türlü metabolit pirüvat dekarboksilasyonu ve
sitrik asit döngüsünü yavaşlatırken, organizmanın ATP açığının
göstergesi olan maddeler de bu olayları hızlandırırlar.
Pirüvatın oksidatif dekarboksilasyonundaki regülatör enzim
pirüvat dehidrogenaz dır. Bu enzim hem allosterik hem de
kovalent modifikasyonla regüle edilir. AMP, Ca2+ ve NAD+ bu
enzimi allosterik olarak aktive ederken, asetil-KoA, ATP , NADH
ve yağ asitleri inhibe ederler. Öte yandan aynı enzimdeki spesifik
serin kökü fosforile olduğu zaman enzim inaktif konuma geçer,
defosforilasyon ise aktiviteyi yeniden kazandırır.
Pirüvat
Dehidrogenazın
kovalent
modifikasyonla
kontrolu
+
Sitrik Asit Döngüsünün Kontrolü
Sitrik asit dögüsünün kontrolünde de aynı temel kural işler. Substratların
varlığı, biriken ürünlerin oluşturduğu inhibisyon ve döngünün son ürünleri
olarak ortaya çıkan metabolitlerin yarattığı allosterik regülasyon bu metabolik
yoldaki kontrolü sağlayan mekanizmalardır. Regülasyon 3 enzim üzerinden
sağlanır.
1. Sitrat sentaz: Bu enzimin kontrol mekanizması iyi bilinmemektedir.
Aktivatör: ADP
İnhibitör: ATP, NADH, süksinil KoA ve sitrat.
2. İzositrat Dehidrogenaz:
Allosterik aktivatör: Ca2+, ADP
Allosterik inhibitör: NADH
E. Coli de kovalent modifikasyonla regülasyon gösterilmiş.
Fosforilasyonla inaktif, defosforilasyonla aktif enzim formu oluşuyor.
3. Alfa-ketoglutarat dehidrogenaz:
Aktivatör: Ca2+
İnhibitör: NADH ve süksinil-KoA nın inhibitör etkisi in vitro olarak
gösterilmiştir.
Pirüvat
dekarboksilasyonu
ve Sitrik asit
Döngüsünün
Kontrolü
H
Sitrik Asit Döngüsünün Metabolik Önemi
Sitrik asit döngüsü birçok metabolik yolun birleştiği bir noktada
yer alır. Bu nedenle karbohidratlardan başka yağlar, amino asitler,
nükleotidler ve porfirinlerin bu metabolik yolla ilişkileri vardır.
Karbohidrat olmayan maddelerden glukoz sentezi de bu
metabolik yolun reaksiyonlarının kullanılması ile gerçekleşir.
Döngüde yer alan ara metabolizma maddeleri, bir katalizör gibi
görev yaparak, çok miktardaki substratın ürünlerine dönmesini
sağlar. Örneğin pirüvatın oksidatif dekarboksilasyonu, yağların
oksidasyonu sonunda meydana gelen asetil- KoA lar katalitik
dozdaki OAA yardımıyla parçalanmaya uğrar. Bu nedenle döngü
ara maddelerinin konsantrasyonlarının korunması önemlidir.
Sitrik Asit Döngüsü ile Metabolik İlişkisi Olan Yollar
Yağ
asitleri
Bir mol glukozun aerobik olarak oksidasyonu
ile elde edilen ATP miktarı
Glikoliz, pirüvatın oksidatif dekarboksilasyonu ve sitrik asit
döngüsünde oluşan ATP miktarları
Glukuronik Asit Metabolik Yolu
Organizmamızda glukozun ufak bir bölümü bu
metabolik yolda kullanılır.
Üronik asit metabolik yolu karaciğerde gerçekleşir
ve yolun başlangıç reaksiyonları glikojen sentezi ile
aynıdır. İlk aşamada glukoz-6-fosfat
fosfoglukomutaz’la glukoz-1-fosfata çevrilir.
Görevleri
Yolun sonunda oluşan UDP
- glukuronat , glukuronik asidin aktif
şekli olup, glukuronik asidin steroid hormonlara, bilirübine, ve
organizmaya yabancı olan bir çok maddeye (ksenobiyotik)
bağlanmasını sağlar. Bu işleme glukuronidasyon denir.
Glukuronik asitle birleşen her madde suda daha iyi çözünür
konuma
gelir
ve
organizmayı
kolayca
terkeder
(detoksifikasyon).
UDP
- glukuronat ayrıca mukopolisakkaridlerin oluşumunda, C
vitamini sentezinde görev alır. Üronik asit metabolik yolu aynı
zamanda glukoz için bir oksidasyon yoludur. Ancak bu sırada
ATP sentezi gerçekleşmez ve oluşan ksilüloz-5- fosfat, pentoz
fosfat metabolik yolu içinde kullanılır.
1. Reaksiyon
Glukoz-6-fosfat, fosfoglukomutaz ile tersinir olarak
glukoz-1-fosfata dönüştürülür.
Glukoz-6-fosfat ↔ Glukoz-1-fosfat
2. Reaksiyon
UDP-glukoz pirofosforilaz glukoz-1-fosfatı UTP ile
birleştirerek UDP-glukoz (üridin difosfat glukoz)
oluşturur.
3. Reaksiyon
UDP-glukoz da UDP-glukoz dehidrogenaz
tarafından 6.karbonundan UDP-glukuronata
oksitlenir.
4. Reaksiyon
UDP-glukuronat; glukuronidasyon dışında, proteoglikan ,
C vitamini ve ksilüloz-5-fosfat sentezlerinde de kullanılır.
Pentoz Fosfat Yolu
Heksozmonofosfat metabolik yolu veya fosfoglukonat yolu
ismi de verilmektedir. Amaç enerji üretimi değil, redükleyici güce
sahip olan NADPH + H+’ları üretmektir. Bu fonksiyon özellikle
karaciğer, adrenal korteks ve meme bezleri gibi yağ asidi ve
steroid sentezi yapan dokuların hücrelerinde önemli bir yer
tutmaktadır.
Heksozların pentozlara dönüsümü: Yolda oluşan Riboz 5- o
f sfat
hem nükleik asit sentezine hem de koenzimlerin koenzimlerin
yapısına girer.
Pentozların oksidatif yıkımları için heksozlara dönüştürülmesinde
kullanılır.
Fotosentezin karanlık devre reaksiyonlarında CO2’den glukoz
oluşumunda görev alırlar.
Pentoz fosfat yolunun oksidatif aşamasında 3 glukoz-6fosfattan 3CO2, 6 NADPH+H+ ve 3 riboz meydana gelir. İkinci
aşama reorganizasyon dönemidir. Burada 3 riboz kendi aralarında
düzenlemeler yaparak sonunda iki glukoz- -6fosfat ve bir
gliseraldehit- -3 fosfat oluştururlar.
GENEL
REAKSİYON
Glukoz 6-fosfat + 2 NAD+ + H2O
2 NADPH + H+ + Riboz 5-fosfat + CO2
1. Reaksiyon
Glukoz- -6 of sfat, 6- fosfoglukonata çevrilirken ilk NADPH + H+
da oluşur. Reaksiyon -6 o
f sfoglukanolakton üzerinden gerçekleşir.
Enzimler: Glukoz-6-fosfat dehidrojenaz ve
6-fosfoglukanolaktonaz
2. Reaksiyon
-6 fosfoglukonat, 6-fosfoglukonat dehidrogenaz
tarafından oksidatif- dekarboksilasyonla ribüloz- -5fosfata
çevrilirken, ikinci NADPH + H+ da sentezlenir ve CO2
açığa çıkar.
3. Reaksiyon - Reorganizasyon
Üç molekül ribüloz- -5 fosfat, ribüloz-5-fosfat epimeraz
ve ribüloz-5-fosfat keto izomeraz ile epimeri olan
ksilüloz- -5 fosfat ve ketoizomeri olan riboz- -5 fosfata
çevrilir.
4. Reaksiyon
Ksilüloz-5-fosfattaki ketol grubu transketolaz ile riboz-5-fosfata
aktarılır ve böylece 7 karbonlu sedoheptüloz -7-fosfat oluşur.
Ksilüloz-5-fosfattan geriye kalan ise gliseraldehit-3-fosfattır.
5. Reaksiyon
Sedoheptüloz- -7 of sfattaki aktif dihidroksiaseton grubu
transaldolaz enzimi ile gliseraldehid- -3 fosfata aktarılır. Bu kez
ürün olarak fruktoz- -6 o
f sfat ve eritroz- -4fosfat ortaya çıkar.
6. Reaksiyon
Transketolaz
reaksiyonu ikinci kez
tekrarlar. Reorganizasyon
için yukarılarda sıra
bekleyen ikinci ksilüloz- -5
fosfatın ketol grubu
eritroz- 4- fosfata aktarılır.
Sonuçta fruktoz- 6- fosfat
ve gliseraldehid
- -3fosfat
oluşur.
Bu şekilde pentoz
fosfat yolunun ikinci
aşaması olan
reorganizasyon
reaksiyonları da
tamamlanmış olur. Bu
kademenin ürünleri
olan iki früktoz-6fosfat ve bir
gliseraldehid-3-fosfat
glikolize girerek
pentoz fosfat yolunun
oluşturduğu by pass
olayını tamamlar.
Pentoz fosfat yolunun
tüm reaksiyonları
şekilde topluca
görülmektedir.