T.C
GAZİ ÜNİVERSİTESİ
SAĞLIK BİLİMLERİ ENSTİTÜSÜ
HİSTOLOJİ VE EMBRİYOLOJİ ANABİLİM DALI
OVULASYON İNDÜKSİYONUNDA OVARYUM PROLİFERASYON
ANTİJENLERİNİN BELİRLENMESİ
YÜKSEK LİSANS TEZİ
Sinem DEMİR
Tez Danışmanı
Prof. Dr. Deniz ERDOĞAN
ANKARA
Ocak 2011
T.C
GAZİ ÜNİVERSİTESİ
SAĞLIK BİLİMLERİ ENSTİTÜSÜ
HİSTOLOJİ VE EMBRİYOLOJİ ANABİLİM DALI
OVULASYON İNDÜKSİYONUNDA OVARYUM PROLİFERASYON
ANTİJENLERİNİN BELİRLENMESİ
YÜKSEK LİSANS TEZİ
Sinem DEMİR
Tez Danışmanı
Prof. Dr. Deniz ERDOĞAN
Bu tez Gazi Üniversitesi Bilimsel Araştırma Projeleri Birimi tarafından 01/2010-47 proje numarası ile
desteklenmiştir.
ANKARA
Ocak 2011
i
İÇİNDEKİLER
Kabul ve Onay
i
İçindekiler
ii
Resimler
iv
Tablolar
x
Kısaltmalar ve Simgeler
xi
1. GİRİŞ
1
2. GENEL BİLGİLER
5
2.1.Ovaryum Embriyolojisi
2.1.1. Oogenezis
2.2. Ovaryum Anatomisi
5
8
12
2.2.1. Ovarium’un Yapısı
13
2.2.2. Ovarium Arterleri
14
2.2.3. Ovarium Venleri
14
2.2.4. Lenf drenajı
15
2.2.5. Ovarium Sinirleri
15
2.3. Ovaryum Histolojisi
2.3.1. Ovaryum Follikülleri
16
17
2.3.1.1. Primordiyal Follikül
17
2.3.1.2. Primer Follikül
18
2.3.1.3. Sekonder Follikül
19
2.3.1.4. Tersiyer Follikül( Graaf Follikül)
20
2.3.1.5. Atretik Follikül
21
2.3.2. Ovulasyon
21
2.3.3. Korpus Luteum
22
2.4. Ovaryum Fizyolojisi
2.4.1.Ovaryum Hormonlarının İşlevi-Östradiol ve Progesteron
24
28
2.4.1.1. Östrojenler
29
2.4.1.2. Östrojenlerin İşlevleri
30
2.4.1.3.Progestinler
32
2.4.1.4. Progesteronların İşlevleri
33
2.4.2. Hipotalamus’tan Salgılanan GnRH ve Bu Hormonun
FH ve LH Salgılanmasına Etkileri
34
ii
2.5. İnfertilite ve Ovulasyon İndüksiyonu
36
2.5.1. İnfertilite
36
2.5.2. Ovulasyon İndüksiyonu
38
2.5.2.1.Gonadotropinler ile Ovulasyon İndüksiyonu
39
2.5.2.1.1.Gonadotropin Endikasyonları
41
2.5.2.1.2.Gonadotropinlerle Tedavi Rejimi
41
2.5.2.1.3.Gonadotropin Tedavi Sonuçları
43
2.5.2.2.Klomifen Sitrat(KS) ile Ovulasyon İndüksiyonu
43
2.5.2.2.1.Klomifen Sitrat Endikasyonları
44
2.5.2.2.2.Klomifen sitrat Dozu
45
2.5.2.2.3.Klomifen Sitrat Tedavi Sonuçları
45
2.5.2.2.4.Klomifen Sitrat’ın Yan Etkileri
46
2.6. Proliferasyon Hücre Çekirdek Antijeni (PCNA)
47
2.7. c-FOS
49
2.8. Lösemi İnhibitör Faktör (LIF)
52
3.GEREÇ VE YÖNTEMLER
54
3.1. Deney Hayvanları ve Gruplandırma
54
3.2. İmmunohistokimyasal Yöntem
56
3.3 İstatistiksel Yöntem
58
4. BULGULAR
59
4.1. PCNA Bulguları
59
4.2. C-FOS Bulguları
61
4.3. LIF Bulguları
62
4.4. İstatistiksel Bulgular
63
5.TARTIŞMA
120
6. SONUÇ
137
7. ÖZET
140
8. SUMMARY
142
9. KAYNAKLAR
144
10.EKLER
160
11.TEŞEKKÜR
161
12.ÖZGEÇMİŞ
162
iii
RESİMLER
Resim 1: Hormon uygulaması yapılan gruplara ait kontrol ovaryum
dokusunda
yapılan
PCNA
boyaması
(İmmünperoksidaz-
HematoksilenX100).
Resim 2: Hormon uygulaması yapılan gruplara ait kontrol ovaryum
dokusunda yapılan PCNA boyaması (İmmünperoksidaz-Hematoksilen
AX400, BX400).
Resim 3: Hormon uygulaması yapılan gruplara ait kontrol ovaryum
dokusunda yapılan PCNA boyaması (İmmünperoksidaz-Hematoksilen
X400).
Resim 4: Hormon uygulaması yapılan gruplara ait ovaryum dokusunda
PCNA
boyaması
(İmmünperoksidaz-Hematoksilen
AX100,
BX100,
CX100).
Resim 5: Hormon uygulaması yapılan gruplara ait ovaryum dokusunda
PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400, BX400).
Resim 6: Hormon uygulaması yapılan gruplara ait ovaryum dokusunda
PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400, BX400).
Resim 7: Hormon uygulaması yapılan gruba ait gebe ovaryumlarında
yapılan PCNA boyaması (İmmünperoksidaz-Hematoksilen AX100, BX100,
CX100, DX100).
Resim 8: Hormon uygulaması yapılan gruba ait gebe ovaryumlarında
PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400, BX400).
Resim 9: Hormon uygulaması yapılan gruba ait gebe ovaryumlarında
PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400).
iv
Resim 10: Klomifen uygulaması yapılan gruba ait kontrol grubu ovaryum
dokusunda yapılan PCNA boyaması (İmmünperoksidaz-Hematoksilen
AX100,BX100).
Resim 11: Klomifen uygulaması yapılan gruba ait kontrol grubu ovaryum
dokusunda yapılan PCNA boyaması (İmmünperoksidaz-Hematoksilen
AX400,BX400).
Resim 12: Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokularında PCNA boyaması (İmmünperoksidaz-Hematoksilen X100).
Resim 13: Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokularında PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400,
BX400).
Resim 14: Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokularında PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400,
BX400).
Resim
15:
Klomifen
sitrat
uygulaması
yapılan
gruba
ait
gebe
ovaryumlarında PCNA boyaması (İmmünperoksidaz-Hematoksilen AX100,
BX100, CX100).
Resim
16:
Klomifen
sitrat
uygulaması
yapılan
gruba
ait
gebe
ovaryumlarında PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400,
BX400, CX400).
Resim
17:
Klomifen
sitrat
uygulaması
yapılan
gruba
ait
gebe
ovaryumlarında PCNA boyaması (İmmünperoksidaz-Hematoksilen AX400,
BX400).
Resim 18: Hormon uygulaması yapılan gruba ait kontrol grubu
ovaryumlarında c-fos boyaması (İmmünperoksidaz-Hematoksilen X100).
v
Resim 19: Hormon uygulaması yapılan gruba ait kontrol grubu
ovaryumlarında c-fos boyaması (İmmünperoksidaz-Hematoksilen AX400,
BX400).
Resim 20: Hormon uygulaması yapılan gruba ait kontrol grubu
ovaryumlarında c-fos boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 21: Hormon uygulaması yapılan gruba ait ovaryum dokusunda cfos boyaması (İmmünperoksidaz-Hematoksilen AX100, BX100).
Resim 22: Hormon uygulaması yapılan gruba ait ovaryum dokusunda cfos boyaması (İmmünperoksidaz-Hematoksilen AX400, BX400).
Resim 23:
Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda c-fos boyaması (İmmünperoksidaz-Hematoksilen AX100,
BX100).
Resim 24: :
Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda c-fos boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 25: Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda c-fos boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 26: Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda c-fos boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 27: Klomifen uygulaması yapılan gruba ait kontrol grubuna ait
ovaryum
dokusunda
yapılan
c-fos
boyaması
(İmmünperoksidaz-
Hematoksilen X100).
Resim 28: Klomifen uygulaması yapılan gruba ait kontrol grubuna ait
ovaryum
dokusunda
yapılan
c-fos
boyaması
(İmmünperoksidaz-
Hematoksilen X400).
vi
Resim 29: Klomifen uygulaması yapılan gruba ait kontrol grubuna ait
ovaryum
dokusunda
yapılan
c-fos
boyaması
(İmmünperoksidaz-
Hematoksilen X400).
Resim 30: Klomifen uygulaması yapılan grup ovaryum dokusunda yapılan
c-fos boyaması (İmmünperoksidaz-Hematoksilen AX100, BX100).
Resim 31: Klomifen uygulaması yapılan grup ovaryum dokusunda yapılan
c-fos boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 32: : Klomifen uygulaması yapılan grup ovaryum dokusunda
yapılan c-fos boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 33: Klomifen uygulaması yapılan gruba ait gebe grubu ovaryum
dokusunda yapılan c-fos boyaması (İmmünperoksidaz-Hematoksilen
AX100, BX100).
Resim 34: Klomifen uygulaması yapılan gruba ait gebe grubu ovaryum
dokusunda yapılan c-fos boyaması (İmmünperoksidaz-Hematoksilen
X400).
Resim 35: Klomifen uygulaması yapılan gruba ait gebe grubu ovaryum
dokusunda yapılan c-fos boyaması (İmmünperoksidaz-Hematoksilen
AX400, BX400).
Resim 36: Hormon uygulaması yapılan gruba ait
kontrol grubuna ait
ovaryum
(İmmünperoksidaz-
dokusunda
yapılan
LIF
boyaması
Hematoksilen X100).
Resim 37: Hormon uygulaması yapılan gruba ait kontrol grubuna ait
ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-
Hematoksilen AX400, BX400).
Resim 38: Hormon uygulaması yapılan gruba ait ovaryum dokusunda
yapılan LIF boyaması (İmmünperoksidaz-Hematoksilen X40).
vii
Resim 39: : Hormon uygulaması yapılan gruba ait ovaryum dokusunda
yapılan LIF boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 40: Hormon uygulaması yapılan gruba ait ovaryum dokusunda
yapılan LIF boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 41: Hormon uygulaması yapılan gruba ait gebe grubu ovaryum
dokusunda yapılan LIF boyaması ). (İmmünperoksidaz-Hematoksilen
X40).
Resim 42: Hormon uygulaması yapılan gruba ait gebe grubu ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-Hematoksilen
AX400, BX400).
Resim 43: Hormon uygulaması yapılan gruba ait gebe grubu ovaryum
dokusunda yapılan LIF boyaması (İmmünperoksidaz-Hematoksilen X400).
Resim 44: Klomifen uygulaması yapılan gruba ait kontrol grubu ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-Hematoksilen
AX100, BX100).
Resim 45: Klomifen uygulaması yapılan gruba ait kontrol grubu ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-Hematoksilen
AX400, BX400, CX400).
Resim 46: Klomifen sitrat uygulaması yapılan gruplara ait ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-Hematoksilen
AX100, BX100).
Resim 47: Klomifen sitrat uygulaması yapılan gruplara ait ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-Hematoksilen
AX400, BX400).
viii
Resim 48: Klomifen sitrat uygulaması yapılan gruba ait gebe grupların
ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-
Hematoksilen AX100, BX100).
Resim 49: : Klomifen sitrat uygulaması yapılan gruba ait gebe grupların
ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-
Hematoksilen X400).
Resim 50: Klomifen sitrat uygulaması yapılan gruba ait gebe grupların
ovaryum
dokusunda
yapılan
LIF
boyaması
(İmmünperoksidaz-
Hematoksilen X400).
ix
TABLOLAR
Tablo 1. Hormon ve klomifen sitrat grubuna ait Graaf değerlerinin
istatistiksel açıdan değerlendirilmesi
Tablo 2. Hormon ve klomifen sitrat uygulaması yapılan gruplara ait
ovaryum çaplarının istatistiksel açıdan değerlendirilmesi
Tablo 3. Hormon ve klomifen sitrat uygulaması yapılan gruplara ait
primordiyal follikül sayılarının istatistiksel açıdan değerlendirilmesi
Tablo 4. Hormon ve klomifen sitrat uygulaması yapılan gruplara ait
sekonder follikül sayılarının istatistiksel açıdan değerlendirilmesi
Tablo 5. Hormon uygulaması yapılan gruba ait tanıtıcı istatistikler
Tablo 6. Klomifen sitrat uygulaması yapılan gruba ait tanıtıcı istatistikler
Tablo 7. Graaf çaplarına ait tanıtıcı istatistikler
Tablo 8. Follikül sayılarına ait tanıtıcı istatistikler
x
KISALTMALAR ve SEMBOLLER
ATP
: Adenozin trifosfat
cAMP
: Siklik adenozin monofosfat
DNA
: Deoksiribonükleik asit
FSH
: Follikül uyaran hormon
GnRH
: Gonadotropin salgılatıcı hormon
hCG
: İnsan koryonik gonadotropin hormonu
hMG
: İnsan menapozal gonadotropin hormonu
KOH
: Denetimli ovaryan hipersitimülasyon
KS
: Klomifen sitrat
LH
: Lüteinize hormon
OHSS
: Ovaryan hiperstimülasyon sendromu
OMI
: Oosit olgunlaşmasını baskılayan madde
PAS
: Periyodik asit Schiff
TBF
: Testis belirleyici faktör
TSH
: Tiroid uyaran hormon
WHO
: Dünya Sağlık Örgütü
ZP
: Zona pellucida
xi
1.GİRİŞ
Ġnfertilite; Dünya Sağlık Örgütü‘ nce (WHO), korunmasız cinsel
yaĢama karĢın bir yıl süresince gebe kalınamaması olarak tanımlanmaktadır.
Üreme yaĢ grubundaki çiftlerin %10-15‘ini etkilemektedir. Ġnfertilite oranı son
yıllarda giderek artmaktadır. Bu artıĢın önemli nedenleri arasında kadınlarda
artan kariyer düzeyi, yüksek evlilik yaĢı ve boĢanma oranı, doğum kontrolü ve
aile planlaması alanında geliĢmeler, gecikmiĢ doğum yaĢı gösterilmektedir.
1,2,3,4
Ġnfertilite
değiĢik
toplumlarda
farklı
nedenlerle
ortaya
çıkmaktadır. Hull ve arkadaĢlarının Ġngiltere Bristol‘da 708 çift arasında
yaptıkları bir araĢtırmada infertilite nedenlerinin %21‘inin ovaryan yetmezlik,
%14‘ünün tubal etken, %21‘inin erkek faktörü ve %28‘inin açıklanamayan
infertilite olduğu bildirilmiĢtir. Günümüzde infertilite nedenleri anovulasyon,
tubal
etkenler,
erkek
faktörü
ve
açıklanamayan
infertilite
olarak
gruplanabilmektedir. Uterin ve endometrial etkenler, gamet ve embriyo
defektleri ve gamet ya da embriyonun mikro çevresini etkileyebilecek
herhangi bir durum da infertilitede rol oynayan diğer faktörlerdir.
5,6
Toplumun büyük bir kesimini ilgilendiren bu sorun, psikolojik ve
sosyal boyutunun yanı sıra son yıllarda ileri düzeyde geliĢme gösteren
yardımcı üreme tekniklerinin de kullanılmaya baĢlanmasıyla, özellikle
geliĢmekte olan ülkeler için büyük finansal sorunları da birlikte getirmektedir.
Bu nedenle infertilite olgularında hastaya uygun sistematik bir yaklaĢımda
bulunulması son derece önemlidir. 7
1
Sorunların aĢılmasına yönelik tedavi yöntemleri arasında
ovulasyon
indüksiyonu
intrauterin
inseminasyon
ile
birlikte
infertilite
tedavisinin temelini oluĢturur. Anovulatuar ya da normal ovulasyonu
gerçekleĢtiren ancak yetersiz folliküler ve luteal geliĢmesi olan hastalarda
infertilite tedavisi ilk seçenek olarak yerini almıĢtır. Ovulasyon indüksiyonu
kapsamında, denetimli ovaryan uyarım için genellikle klomifen sitrat kullanılır.
Klomifen sitrat‘ın infertilite tedavisinde ilk aĢama olarak seçilmesinin nedeni
diğer tedavi seçeneklerine karĢın ciddi yan etkisi az, maliyeti düĢük, baĢarısı
yüksek bir tedavi seçeneği olmasından kaynaklanmaktadır. Klomifen sitrat
kimyasal olarak bir trifeniletilen türevidir. Enklomifen ve zuklomifen olarak iki
izomerin
karıĢımından
oluĢmaktadır.
Enklomifen
daha
çok
oranda
antiöstrojenik erke sahiptir. Klomifen sitrat, hipofiz ve hipotalamusta östrojen
reseptörlerini bloke ederek bu merkezlerin östrojenlerin
olumlu geri
denetimlerinden kurtulmasını sağlar. Klomifen sitrat östrojen reseptörlerine
bağlandığında bu reseptörleri birkaç haftaya varan sürelerde iĢgal eder.
7,8,9,10
Klomifen sitrat tedavisine yanıt vermeyen hastalara bir üst
aĢama
olarak,
―insan
menopozal
gonadotropin‖
(hMG)
ile
tedavi
denenmektedir. FSH ve LH‘nın granuloza ve teka hücreleri üzerindeki etkileri
çok iyi belirlenmiĢtir ve folliküler geliĢim için esastır. Etki düzeneği klomifen
sitrata karĢın daha iyi bilinmektedir. Tedavinin baĢarılı olabilmesi için
ovaryumların
iĢlevsel
gonadotropinlerin
olması
yerini
alarak
gerekir,
çünkü
ovulasyon
verilen
indüksiyonu
hMG
endojen
yapmaktadır.
Kendiliğinden olan döngülerde olduğu gibi FSH birincil olarak granüloza
hücreleri ve LH teka lutein hücreleri üzerinde etki göstererek folikülogenezisi
uyarırlar. 11,12
2
Ġnsan koryonik gonadotropin (hCG) ise, karbonhidrat yan zincirli
glikoprotein yapılı bir hormondur. LH, FSH ve TSH‘nın heterodimerik yapısı
ile
benzerlik
gösterir.
Mazer
ve
Ravetz
gebe
kısrakların
serum
gonodotropinlerini ve koyunların hipofiz ekstrelerini follikül uyarımı için ve
hCG‘yi folikül olgunlaĢmasının sonlandırılması için amenoreik kadınlarda
kullanmıĢlar ve hipofizer bir faktörün folikül geliĢimini uyardığı ve hCG‘nin
ovulasyonu indüklediği sonucuna varmıĢlardır.
13,14
Bugün ovulasyon indüksiyonunda amaç, anovulatuar hastalarda
ovulasyonu baĢlatmaktır. Ovulasyon sağlanabildiği durumlarda ise esas amaç
tek folliküllü ovulasyondur. 15
Ġnfertilite
tedavisinde
elde
edilen
baĢarıya
koĢut
olarak
ovulasyon indüksiyon ajanlarının kullanımı her geçen gün artmaktadır. Bu
artıĢ aynı zamanda istenmeyen yan etkileri de beraberinde getirmektedir. En
sık rastlanan ciddi yan etki ovaryan hiperstimulasyon sendromu (OHSS)‘dur.
Bu uyarım protokollerinin % 0.3-5‘inde oluĢan ölümcül olabilecek bir
durumdur. Ovulasyon indüksiyonunda %35 oranında karĢılaĢılan çoğul
gebelikler beraberinde getirdiği morbidite ve fötal mortalite nedeniyle
istenmeyen durumlardan biridir. Ayrıca ovulasyon indüksiyonu sonrası meme,
ovaryum ve endometriyum kanseri dağılımında artıĢ ile ilgili çok sayıda olgu
sunumları bildirilmiĢtir. 6,16
Bu konuyla ilgili olarak yapılan diğer çalıĢmalarda ise üst üste
yapılan hormonal ovulasyon induksiyonu uygulamalarının, önlenemez hücre
çoğalmasına ve kansere neden olduğu da bildirilmiĢtir. Tüm bu çalıĢmalara
ek olarak bu maddelerin kullanımının ovaryumda tam olarak ne gibi histolojik
değiĢikliklere neden olduğu açıklığa kavuĢmamıĢtır. 17
3
Bizim
çalıĢmamızda,
klomifen
sitrat ve
gonodotropinlerin
ovulasyon induksiyonu için kullanımları sonucunda ovaryumlarda neden
olabileceği yapısal değiĢimlerin immunohistokimyasal olarak karĢılaĢtırmalı
incelenmesi amaçlanmıĢtır. Bu amaçla hücre proliferasyon iĢaretleyicileri olan
PCNA, LIF ve c-fos belirteçleri kullanılmıĢ ve bulgular karĢılaĢtırmalı olarak
değerlendirilmiĢtir.
4
2.GENEL BİLGİLER
2.1.OVARYUM EMBRİYOLOJİSİ
Embriyonun
cinsiyeti,
genetik
olarak
döllenme
sırasında
belirlenmiĢ olmasına karĢın, gonadlar geliĢimin 7. Haftasına değin erkek ya
da diĢi yapısal özelliğe sahip değillerdir. 18
Cinsiyetin farklanması çok sayıda genin rol oynadığı karmaĢık
bir süreçtir. SRY geni (Y kromozomu ‗nun üzerindeki cinsiyet belirleyici bölge)
Y kromozomunun kısa kolunda (Yp11) bulunur ve cinsiyet belirleyicisidir. Bu
genin protein ürünü olan transkripsiyon faktörü, geliĢmemiĢ durumdaki üreme
organlarının cinsiyetini belirleyen genleri harekete geçirir. SRY geni Testis
Belirleyici Faktör‘dür(TBF). Bu faktörün var olması durumunda fetusun
cinsiyeti erkek, yokluğunda ise kızdır. 18
Gonadlar baĢlangıçta kölom epitelinin çoğalması ve altındaki
mezenĢimin yoğunlaĢmasıyla oluĢmuĢ bir çift genital ya da gonadal kabartılar
Ģeklinde belirirler. GeliĢimin 6. haftasına değin bu kabartılar içinde üreme
hücreleri görülmez. 18,19
Ġlkel (primordiyal) üreme hücreleri, geliĢimin 2. haftasında primer
embriyonik ektodermden (epiblast) köken alırlar. Daha sonra epiblasttan
ayrılarak ameboid hareketlerle vitellus kesesinin ekstraembiyonik yapılarına
5
göç ederler. BaĢlangıçta embriyonun kaudalinde yerleĢik ekstraembriyonik
mezodermde görülürler, sonra vitellus kesesi duvarı endoderminde izlenirler.
GeliĢimin 4. haftasında son bağırsağın mezenterinin dorsali boyunca
ilerleyerek 5. hafta baĢında ilkel gonadlara ulaĢırlar. 6. haftada ise genital
kabartılara yerleĢirler. Ġlkel üreme hücrelerinin izlediği bu yola germ çizgisi
(hattı) denir. Ġlkel üreme hücreleri açık renk sitoplazmaları ve oval Ģekilleriyle
ayırt edilirler. Bu hücreler genital kabartılara ulaĢamadıklarında gonadlar
geliĢemez. Gonadların ovaryum ya da testis‘e farklanmalarında ilkel üreme
hücrelerinin tetikleyici etkisi vardır. 18,19
Ġlkel üreme hücrelerinin ilkel gonadlara ulaĢmasından hemen
önce ve ulaĢması sırasında, genital kabartının epiteli çoğalır ve epitel
hücreleri altındaki mezenĢimin içine gömülürler. Burada ilkel cinsiyet
kordonları olarak adlandırılan düzensiz Ģekilli kordonları oluĢtururlar. Erkek ve
diĢi embriyolarda bu kordonlar yüzey epiteline bağlıdır ve bu evrede erkek ya
da diĢi gonadlarının birbirinden ayırt edilmesi olanaksızdır. Bu nedenle bu
evreye farklanmamıĢ evre, bu gonada da farklanmamıĢ gonad denir. 18
XX
10.haftaya
değin
kromozomları
ovaryumlar
ovaryumun
histolojik
geliĢimi
olarak
için
ayırt
genler
içerir.
edilmezler.
XX
kromozomlarına sahip diĢi embriyolarda, ilkel cinsiyet kordonları düzensiz
hücre kümelerine ayrılır. Ġlkel üreme hücresi grupları içeren bu kümeler, daha
çok ovaryum‘un medullar bölgesinde yerleĢmiĢlerdir. Bu hücre kümeleri bir
süre sonra kaybolarak yerlerini damarlı stromaya (medulla) bırakırlar.
18,19
6
DiĢi gonadın yüzey epiteli, erkek gonadın aksine çoğalmayı
sürdürür. 7. haftada yüzey epitelinden köken alan hücre kordonları alttaki
mezenĢim içine gömülür. Bunlar yüzeye yakın yerleĢirler ve ikincil kordonlar
olarak adlandırılırlar. 4. ayda, bu kordonlar bir ya da daha çok sayıda ilkel
üreme hücresini çevreleyen izole hücre toplulukları haline gelirler. Bu üreme
hücreleri zamanla oogonyumlara dönüĢürken yüzey epitelinden aĢağıya göç
eden ve üreme hücrelerini çevreleyen epitel hücrelerinden de foliküler
hücreler oluĢur. 18,19
Germ hücreleri göçleri sırasında bir yandan da çoğalırlar. Bunlar
spermiyum ve oositlerin öncülleridir. 6. haftanın sonunda mitoz bölünmeyle
çoğalan hücrelerin sayıları kadardır. 20,21
Ovaryum farklılaĢmasının ilk belirtileri 6 ile 8. haftalar arasında
germ hücrelerinin hızla mitotik bölünmesidir. Bu bölünme sonucunda 16 ile
20. haftalar arasında yaklaĢık 6-7 milyon oogonyum oluĢur. Bu evreden
baĢlayarak ovaryum‘un üreme hücre sayısı geri dönüĢümsüz azalır ve
yaklaĢık 50 yıl sonra da tüm oosit içeriği tükenir. 20‘li yaĢlarda 104.000, erken
30‘lu yaĢlarda 33.000 düzeyine inen folikül ve oosit sayısı 40‘lı yaĢlarda
7900‘e kadar düĢer. 21
7
2.1.1.Oogenezis
Ġlkel üreme hücreleri, diĢi gonada ulaĢır ulaĢmaz genetik olarak
oogonyumlara farklanırlar. Ardı ardına geçirdikleri mitotik bölünmelerle
çoğalan oogonyumlar 3. ayın sonunda kümeler halinde dizilir, yassı epitel
hücreleriyle çevrelenirler. Yassı epitel hücreleri follikül hücreleri olarak
isimlendirilirler ve ovaryum yüzey epitelinden köken alırlar. Bir küme içindeki
oogonyumların tümü, tek bir ilkel üreme hücresinden geliĢir. 18
Oogonyumların çoğunluğu mitozla bölünmeyi sürdürürken, bir
kısmı bölünmesini I. Mayoz bölünmenin profaz evresinde durdurarak primer
oositlere farklanır. Bundan sonraki birkaç ay içinde oogonyumların sayısı
hızla artar. GeliĢimin 5. ayında ovaryum içindeki germ hücrelerinin sayısı
ortalama 7 milyona ulaĢır. Bu evreden sonra hücre ölümü baĢlar. Çok sayıda
oogonyum ve primer oosit atretik hale gelir. 7. ayda yüzeye yakın yerleĢmiĢ
birkaç oogonyum dıĢında çoğu dejenere olur. Hayatta kalan primer oositlerin
tümü I. Mayoz bölünmenin profaz evresine girerler ve her biri tek sıra yassı
folikül hücre katmanıyla çevrelenirler. Primer oosit, çevresindeki yassı epitel
hücreleriyle birlikte primordiyal follikül adını alır. 18,19
Doğuma yakın evrede primer oositlerin tümü I. Mayoz
bölünmenin profazının diploten evresindedirler. Primer oositler puberteye
değin
dinlenme
evresinde
kalırlar
ve
birinci
mayoz
bölünmelerini
tamamlamazlar. Bu süre boyunca oosit olgunlaĢması folikül hücrelerince
salgılanan oosit olgunlaĢmasını baskılayan madde (OMI) ile baskılanır.
Doğumda ovaryumlardaki primer oosit sayısı 700 bin ile 2 milyon arasında
8
değiĢir. Puberteye değin bunların büyük çoğunluğu atretik hale geldiğinden,
puberte baĢlangıcındaki bu sayı 400.000‘ e düĢer ve 500‘den daha azı bir
kadının üreme döngüsü içinde ovulasyonla atılır. Diğerleri çeĢitli geliĢim
evrelerinde dejenere olup atretik hale gelir. Geç olgunlaĢan oositlerin bazıları
40 yıl süresince I. Mayoz bölünmenin profaz evresinde bekler.
18,19,21
Puberteye gelindiğinde diĢi gonad primordiyal foliküllerden
beslenen bir folikül havuzuna sahip olmuĢ durumdadır. Her ovaryal döngüde
bu havuzdan seçilen 5 ile 15 arasında değiĢen sayıda primordiyal folikül,
primer (preantral), sekonder (antral) ve Graaf ( tersiyer, preovulatuvar) olarak
adlandırılan 3 aĢamadan geçerek olgunlaĢmaya baĢlar. Bunlar arasında en
uzun olanı antral folikül evresidir. Preovulatuvar evre yaklaĢık 37 saat sürer. I.
Mayoz bölünmenin diploten evresinde dinlenme aĢamasında olan primer
oosit büyümeye baĢlar. Bekleme evresinden çıkan bu folliküllere primer
follikül denir ve bunlar iki tiptir:
1.
Tek katmanlı primer folliküller: Bunların çevresinde
tek sıralı kübik folliküler hücreler bulunur.
2.
Çok katmanlı primer folliküller: Çok sıralı follikül
hücre katmanıyla çevrilidirler. Folliküler hücreler, kendilerini ovaryum
stromasından ayıran bir bazal lamina ile desteklenir.
Primer follikül evresinde primer oosit zona pellusida denilen
glikoprotein bir kılıf sentezlemeye baĢlar. Zona pellusida folikül hücrelerini
oositten ayırır. Folliküler hücrelerin ince sitoplazmik uzantıları zona
9
pellusidayı deler ve oositin mikrovilluslarıyla iliĢki kurar. ĠliĢki bölgelerinde
oluklu bağlantılar bulunur. 18,19,22
Bunu izleyen evrede follikül hücreleri sürekli olarak çoğalıp çok
katmanlı follikül hücrelerini oluĢtururlar. Zona pellusida kalınlaĢır. Follikülü
çevreleyen stromal hücreler, teka( kılıf) denilen hücresel bir kılıf oluĢturulacak
Ģekilde düzenlenirler. Teka katmanı daha sonra iki tabakaya farklanır: (1)
Teka interna (iç kılıf) (2) Teka eksterna (dıĢ kılıf). GeliĢmekte olan folikülün
bazal laminasına komĢu, iyi damarlanmıĢ teka interna katmanı hücreleri
steroit salgı oluĢturur. Teka eksterna ise kapsül benzeri bir bağ dokusu
katmanıdır ve ovaryum‘un stroması ile karıĢır. Bu hale gelen follikül sekonder
folliküldür. 22
Folliküler hücreler arasında, küçük hücreler arası boĢluklar CallEksner cisimcikleri görülür. Bu aralıklar folliküler sıvı içerir ve daha sonra
birleĢerek, daha büyük bir boĢluk olan follikül antrumunu yapar. Antrumun
oluĢmasıyla, foliküler hücreleri primer oosit çevresinde yeniden düzenlenirler.
Oosit ile folikül duvarı arasında bulunan bir grup foliküler hücre kümesine
kumulus ooforus denir. En büyük follikül, olgun folliküldür. Buna Graaf follikülü
ya da preovulatuvar folikül denir. 15- 20 mm çapındadır. Ovulasyondan
hemen önce primer oosit, follikül içinde eksentrik bir konum alır. Primer oosit
zona pellusidaya sıkıca yapıĢmıĢ ve korona radiata denilen tek sıralı kübik ya
da prizmatik foliküler hücrelerle çevrilidir. 22
Ovulasyondan birkaç saat önce I. Mayoz bölünme tamamlanır.
Bunun sonucunda sekonder oosit ve birinci kutup cismi olĢur. Birinci kutup
10
cismi perivitellin aralık denilen zona pellusida ile oosit arasındaki aralığa atılır.
I. Mayoz bölünmenin tamamlanmasından hemen sonra ve sekonder oositin
çekirdeği dinlenme evresine geçmeden önce, DNA eĢleĢmesi olmaksızın
hücre II. Mayoz bölünmeye girer. II. Mayoz bölünme ancak oosit II bir
spermiyumla döllenirse tamamlanır. Döllenme olmazsa ovulasyondan 24 saat
sonra oosit II dejenere olur. Ovaryumda oogonyumlardan oosit II oluĢuncaya
kadar gerçekleĢen değiĢimlere oogenezis denir.
18,19,22
11
2.2.OVARYUM ANATOMİSİ
Ovariumlar; oval biçimde bademe benzer bir çift organdır.
Erkekte testisin eĢidir. Her biri 5-6 gr ağırlığa, 3-3.5 cm boya, yaklaĢık 1 cm
ene sahiptir. Sağda ve solda, pelvis duvarında fossa ovarica denilen
çukurlara yerleĢiktirler. Bu çukurlar a.iliaca externa ve a.iliaca interna
arasındadır. Bu çukuru alt- ön taraftan lig. latum uteri tabanı, yukarıda a.
iliaca externa ve arkadan ureterler ile sınırlar. Çukurun dibinde ve peritonun
altından a.-v. Obturatoriale n. Obturatorius geçer. Ġlk gebelikte ovariumlar
uterus ile karın boĢluğuna doğru çekilirler ve bir daha aynı yerlerine
dönemezler. 23,24
Tuba uterina‘nın arka ve alt kısmında bulunan ovaryumlar lig.
Latum uteri içinde bulunurlar. Pembemsi-gri renkli olan ovaryumların yüzü
puberteye değin peritonla örtülü olup düz ve parlaktır. Puberteden sonra
periton özelliğini yitirir ve matlaĢır. Ovulasyon ve doğurmaya bağlı olarak
üzeri pürtüklü bir görünüm alır. 23
Facies lateralis ve facies medialis olarak iki yüzü, extremitas
tubaria ve extremitas uuterina olarak iki ucu ile margo liber ve margo
mesovaricus olmak üzere iki kenarı vardır. 23
V. iliaca externa ‗ya komĢu olan extremitas tubaria‘ ya fimbria
ovarica ve lig. Ovarii suspensorium tutunur. Lig. Suspensorium ovarii, iliak
damarların ön tarafında yukarı doğru uzanan bir periton plikası olup içinde a.
12
ve v. ovarica bulunur. Extremitas uterina aĢağı pelvis döĢemesine doğru
yönelmiĢtir. Bu uç lig. ovarii proprium aracılığı ile uterus‘un cornu uteri‗sine
tutunur. 23
Facies lateralis, fossa ovarica‘yı örten parietal peritona oturur.
Facies medialis dıĢ yüze oranla daha konvekstir. Bu yüzü infindibulum tubae
uterinae örter. Arka kenarına serbest olması nedeniyle margo liber denilir. Ön
kenara oranla daha konveks ve künt olan bu kenar, ureter ile komĢuluk yapar.
Ön kenarına mesovarium tutunması nedeni ile margo mesovaricus denilir.
Arka kenara oranla daha ince olan ön kenarda hilum ovarii bulunur ve a.
umbricalis‘ in artığı ile komĢudur. Bu kenara lig.latum uteri‗nin bir bölümü olan
mesovarium tutunur. Mesovarium‗un iki yaprağı arasında bulunan damar ve
sinirler, hilum ovarii‘den ovarium‘a girer ve çıkarlar. Tuba uterina önce margo
mesovaricus üzerinden kavisle geçer. Daha sonra extremitas tubaria‘nın
üzerinden kıvrılarak margo liber‘e ve buradan da facies medialis‘e gelir.
23
2.2.1.Ovarium’un yapısı
Ovarium en dıĢtan periton ile örtülüdür. Periton tek sıralı yassı
epitel yapısında, düz ve parlaktır. Puberteden sonra ovarium‘un peritonu
yapısını değiĢtirir ve germinal epitel (Waldeyer tabakası) denilen tek sıralı
prizmatik hücre katmanı ile kaplanır. Bu nedenle de mat-gri renge döner.
Epitel katmanının altında tunika albuginea denilen kalın bir tabaka bulunur.
Bu tabaka yaĢ ilerledikçe kalınlaĢır. Tunika albuginea‘nın altında ovarium‘un
esas yapısını oluĢturan stroma ovarii bulunur. Stroma ovarii de, cortex ovarii
ve medulla ovarii denilen iki farklı bölümden oluĢur. Cortex ovarii dıĢ tarafta
13
bulunur ve hilum ovarii dıĢında ovarium‘un her tarafını bir kabuk gibi sarar. Bu
bölümde bağ dokusu arasında değiĢik geliĢme evrelerinde ovarium follikülleri
bulunur. Medulla ovarii iç tarafta yerleĢiktir ve cortex ovarii‘ye oranla daha
gevĢek bir yapıya sahiptir. Medulla ovari‘den gelen bağ dokusu uzantıları
hilum ovarii‘de damarların etrafındaki bağ dokusu ile devam eder. Hilum
ovarii‘den içeri giren damarların çoğu cortex ovarii‘de dağılır. Burada geliĢen
folliküller çevresinde kapiller ağlar oluĢtururlar. 23
2.2.2. Ovarium Arterleri :
Abdominal aorta‘dan çıkan a. ovarica‘lardır. A. ovarica lig.
suspensorium ovarii içinde pelvise iner. Hilum ovarii‘den ovarium‘a girer ve
folliküller çevresinde kılcal ağlar oluĢturur.
2.2.3. Ovarium Venleri:
Arterleri izleyerek hilum ovarii‘den çıkar. Bu venler plexus
pampiniformis denilen venöz ağı oluĢtururlar. Bu ağı oluĢturan venler yukarı
çıktıkça birbirleriyle birleĢirler ve sonunda v. ovarica‘yı oluĢtururlar. V. ovarica,
a.ovarica ile birlikte seyreder. Sol tarafı v. renalis‘e , sağ tarafı ise v.cava
inferior‘a açılır.
14
2.2.4.Lenf drenajı:
Kan damarları ile birlikte uzanır ve nodi lymphatici preaortici ve
nodi lymphatici aortici lateralis‘lere açılırlar.
2.2.5.Ovarium Sinirleri:
Plexus hypogastricus inferior (plexus pelvicus) ve a.ovarica‘nın
çevresindeki plexus ovaricus‘dan gelir. Parasempatikleri n. Vagus‘tan,
sempatikleri ise n. splanchnicus minor ve bir kısım torakal medulla spinalis
segmentlerinden gelir. Bunların ovarium içindeki dağılıĢı ve görevleri tam
olarak bilinmemektedir. 23
15
2.3.OVARYUM HİSTOLOJİSİ
Ovaryum tek katlı yassı epitelden alçak boylu kübik epitele
değin değiĢkenlik gösteren bir epitel ve epitelin hemen altındaki tunika
albugina denilen bir bağ dokusu katmanıyla çevrelenmiĢtir. Enine kesitlerde
dıĢta korteks (kabuk), içte medulla (öz) birbirinden kesin bir çizgiyle ayrılmaz.
Korteks katmanı oldukça kalındır. Bağ dokusu ve çeĢitli geliĢim evrelerinde
follikülleri içerir. 22
Medulla kollajen demetlerden ve elastik lif ağlarından zengin sıkı
bağ dokusudur. Medullada kan ve lenf damarları, kalın sinir demetleri ve hilus
yakınında birkaç düz kas hücresi izlenir. 24
Korteks stroma ve stromada yerleĢik çeĢitli geliĢim evrelerindeki
ovaryum folliküllerinden oluĢur. Stroma kollajen ve elastik lifler ile retiküler lif
ağları ve iğ biçimli bağ dokusu hücrelerinden yapılmıĢtır. Korteksin bu
intersitisyal bağ dokusu, germinal epitel altında sıklaĢarak tunika albuginea‘yı
oluĢturur. 24
Kortekste stroma içinde çeĢitli geliĢim evrelerinde folliküller
yerleĢmiĢtir. Ovaryum follikülleri ortada iri bir germ hücresi (oosit) ile bunu
çevreleyen follikül epitelinden oluĢan yapılardır. 24
16
2.3.1.Ovaryum follikülleri:
Yenidoğan bir çocuğun her iki ovaryumunda toplam 400.000
adet follikül bulunur. Bunlar 40 mikron çapında yuvarlak oluĢumlardır ve iri bir
oosit ile bunu çevreleyen yassı follikül epitelinden oluĢmuĢtur. Bunlara
primordiyal folliküller denir. Her 28 günde bir ovaryumlardan bir oosit atılır. Bir
kadının üreme süreci 30-40 yıl sürer. Bu süre içerisinde yaklaĢık 450 oosit
atılır. 24
2.3.1.1.Primordiyal Follikül
Folliküllerin geliĢiminde ilk fiziksel belirtiler oositin büyümesi ve
granüloza hücrelerinin görünümündeki değiĢikliklerdir. Primordiyal folliküller
fötal yaĢam sırasında oluĢan tek sıralı yassı follikül hücreleriyle çevrili primer
oositleri içerir. Primer oositler organellerden zengindirler. Çok sayıda
mitokondriyon, geliĢmiĢ Golgi kompleksi, granüllü endoplazmik retikulum ve
birkaç ribozom içerirler. Primordiyal folliküller korteksin en üst katmanında yer
alırlar. Sayıları artmaz ancak büyüyüp geliĢirler ve çoğu da dejenere olur.
25,26,27
17
2.3.1.2.Primer Follikül
Pubertenin baĢlamasıyla, her menstrual döngüde 5- 15 arası
primordiyal follikül olgunlaĢmaya baĢlar. Büyümenin ilk iĢareti yassı follikül
epitel hücrelerinin kübikleĢmesidir. Bu aĢamada follikül tek tabakalı
(unilaminar) primer follikül adını alır. Ġlerleyen aĢamalarda follikül hücrelerinin
çoğalmasıyla çok tabakalı (multilaminar) primer follikül oluĢur. Çoğalan follikül
hücreleri de granüloza hücreleri adını alır. Granüloza hücreleri ile oosit
arasında oluklu bağlantılar (nekzus) oluĢur. Bunlar besleyici maddelerin,
iyonların ve düzenleyici moleküllerin geçiĢine olanak sağlar. 24,25,26,27
Granüloza hücreleri çoğalırken çevredeki stroma hücreleri de
follikülü bağ dokusundan oluĢmuĢ bir kılıfla sararak teka follikülü katmanını
oluĢturur. Teka katmanı geliĢtikçe iki tabakaya ayrılır. içte teka interna denilen
oldukça damarlı, kübik salgı hücrelerinden oluĢan kat yer alır. Bu kübik
hücreler
farklılaĢarak
steroid
hormon
üreten
hücrelere
dönüĢürler.
LuteinleĢtirici hormon (LH) reseptörü taĢıyan hücreler östrojen öncülü
androjenleri salgılarlar. Teka interna katmanı fibroblastlar, kollajen lifler ve
küçük damarlardan oluĢur. DıĢta bulunan diğer kat teka eksternadır. Ġçerdiği
kollajen demetler ve düz kaslarıyla daha çok bir kapsül görevi görür. Teka
eksterna katmanı ile ovaryum‘un stroması arasındaki sınır tam olarak belirgin
olmamasına karĢın teka interna ile granüloza katmanı arasındaki sınır burada
bulunan bazal lamina (membrana limitans eksterna) ile oldukça iyi
belirlenmiĢtir. 27
18
Oosit,
follikül
geliĢimi
sırasında
kendi
çevresinde,
glikozaminoglikan ve glikoproteinlerden zengin bir katman olan zona
pellusidayı (ZP) oluĢturur. ZP, homojen, asidofilik, PAS (+) ve oldukça iyi
boyanan jel kıvamında bir yapıdır. Zona pellusida ZP1, ZP2 ve ZP3 denilen 3
farklı glikoproteinden oluĢur. 27
2.3.1.3. Sekonder follikül
Büyümeyi sürdüren follikül içinde sıvı dolu boĢluklar (antrum)
oluĢur. Kısa sürede boĢluklar birbirleriyle birleĢerek yarım ay Ģeklinde follikül
antrumunu Ģekillendirir. Folliküler boĢluğu dolduran sıvı kan damarlarından
plazmanın sızmasıyla oluĢup yerel salgıların metabolik ürünleri ile değiĢmiĢ
hale gelir. BoĢluk oluĢtuktan sonra follikül sekonder (antral) follikül adını alır.
22,26,27
Sekonder follikülün granüloza hücrelerinin çoğalması hipofiz
bezinin ön lobundaki bazofil hücrelerinden salınan FSH ‗a bağlıdır. FSH‘ ın
etkisi altında granüloza hücrelerinin sayısı artar ve hücrelerarası boĢluğa
follikül sıvısı birikir. Bu sıvı glikozaminoglikan, proteoglikan ve hormon
bağlayıcı
proteinler
içeren
kan
plazması
benzeridir.
Follikül
sıvısı
hormonlardan progesteron, östradiol, inhibin, follistatin ve FSH ile LH
salınımını düzenleyen aktivin içerir. 26,27
19
Granüloza katmanı oositi çevreleyen bölge dıĢında her yerde
aynı kalınlıktadır. Antrumda kenarda yerleĢik oosit çevresinde ise granüloza
hücreleri daha yoğun bir Ģekilde birikerek bir tepecik oluĢturur. Buraya
kumulus ooforus denir. Buradaki granüloza hücreleri ovulasyon sonrasına
değin oosit ile birleĢik halde kalırlar. Zona pellusida çevresinde gevĢek
yerleĢmiĢ kübik ya da alçak boylu prizmatik granüloza hücreleri sitoplazmik
uzantılarıyla oosit ile bağlantılarını sürdürür. Primer oositi çevreleyen bu tek
katlı granüloza hücre katmanına korona radiata denir. Oosit sitoplazmasında
mitokondriyonlar çekirdek yakınında toplanırlar. Sitoplazmada protein ve lipit
yapısında vitellus oluĢmaya baĢlar. Oosit 60-80 mikron büyüklüğe ulaĢır.
24,26,27
2.3.1.4.Tersiyer Follikül ( Graaf follikülü)
Graaf follikülünde granüloza hücreleri çoğalır, antrum geniĢler.
Follikül boĢluğu follikül sıvısı ile dolar. Oositi saran zona pellusida‗ya komĢu
ilk sıra granuloza hücreleri tek katlı kübik ya da prizmatiktir. Bunun
dıĢındakiler poligonal biçimlidir. Graaf follikülünü en dıĢtan saran bazal
membran kalın, homojen ve saydamdır. Buna camsı zar denir. Tersiyer
follikül döngünün baĢlangıcından baĢlayarak 10-12 günde oluĢur. 24,26,27
20
2.3.1.5.Atretik Follikül
Çok sayıda primer follikül olgunlaĢma sürecine baĢlasada
bunlardan yalnızca bir tanesi geliĢimini tamamlar, diğerleri atrezi denilen bir
olayla dejenere olur. Bu nedenle ovaryum folliküllerinin büyük bölümü follikül
hücreleri ve oositin ölümüyle geriler ve fagositoz yapan hücrelerce ortadan
kaldırılır. Folliküller geliĢimlerinin herhangi bir evresinde atreziye uğrayabilir.
Atretik folliküller çok değiĢikliğe uğramamıĢ zona pellusida, dejenere olmuĢ
oosit ve follikül hücre kalıntıları ve makrofajlar ile ayırt edilir. Atretik folliküller
doğum öncesinden baĢlayarak menapozun birkaç yıl sonrasına kadar
oluĢurlar. 22
2.3.2.Ovulasyon
Ovulasyondan yaklaĢık 24 saat önce adenohipofizden LH
salınımı tetiklenir. Kandaki LH hormonu düzeyinde oluĢan aĢırı yükselme
sonucu granüloza hücrelerindeki LH reseptörleri duyarlılığını yitirir ve LH‘a
yanıt olarak daha fazla östrojen üretemezler. Oositin I. Mayoz bölünmesi bu
tetiklenme ile durakladığı yerden devam etmeye baĢlar. LH yükselmesinden
12-24 saat kadar sonra gerçekleĢen bu olay, oositin mayoz I ‗i tamamlayarak
ilk kutup cisimciğini oluĢturmasını ve aynı zamanda II. Mayoz bölünmenin
baĢlamasını sağlar. Ancak oosit ovulasyondan yaklaĢık 3 saat öncesine
değin metafaz evresinde duraklar. Bu sırada ovaryum‘un yüzeyinde bir
kabarıklık oluĢur. Bu kabarıklığın tam tepesinde stigma adı verilen damarsız
bir nokta belirir. Yüksek LH düzeyi kollajenaz aktivitesini arttırır ve follikülün
çevresindeki kollajen lifleri sindirir. LH düzeyindeki ani artıĢın diğer bir etkisi
21
de prostaglandin miktarını arttırmasıdır. Prostaglandin ovaryum duvarında
yerel kas kasılmalarına neden olur. Bu kasılmalarla olgun follikül (Graaf
follikülü) yırtılarak içerdiği oositi tuba uterinanın fimbriyaları yakınında periton
boĢluğuna atar. Fimbriyaların hareketi ile oosit tuba uterina içine alınır.
22,24,27,28
Ovulasyon puberteden menapoza değin 28 günde bir olaylanır.
Ovulasyon iki menstrual kanama arasındaki döngünün tam ortasında olur.
Ovulasyon döngünün ortalama 14. gününde gerçekleĢir. Ovulasyonda
genelde bir oosit atılır. Bazen aynı anda birkaç oosit atılabilir. Bu da ikiz ya da
çoklu gebeliklere neden olur. 24
2.3.3. Korpus Luteum
Ovulasyondan sonra granüloza ve teka interna hücreleri
bölünmeden büyüyerek hormon salgılayan lutein hücrelerine dönüĢürler.
Böylece geçici bir endokrin bez olan korpus luteum (sarı cisim) oluĢur. Korpus
luteum progesteron ve östrojen hormonları salgılar. Graaf follikülünde
ovulasyon sonrası bir iç basınç düĢmesi olur ve teka katmanındaki kan
damarlarından kan sızması ile kısa bir süre için follikül içine kan toplanır.
Buna kırmızı cisim ( korpus rubrum, korpus hemorajikum) denir.
24
Döllenme olmadığında, ovulasyonu izleyen 9. günde korpus
luteum en fazla eriĢebileceği boyuta ulaĢır ve ovaryum‘un yüzeyinde
sarımtrak
bir
çıkıntı
halinde
izlenir.
Daha
sonra
luteal
hücrelerin
22
dejenerasyonuyla giderek küçülür ve korpus albikans olarak bilinen fibrotik
skar dokusu haline gelir. Bu sırada progesteron üretimi de azaldığından
menstrual kanama baĢlar. Oositin döllenmesi durumunda korpus luteumun
dejenere olması geliĢmekte olan embriyonun sinsityotrofoblastları tarafından
salgılanan koriyonik gonodotropin ( hCG) hormonu tarafından engellenir.
Böylece korpus luteum büyümesini sürdürür ve gebelik korpus luteumu‘na
dönüĢür. Bu yapı 3. ayın sonunda ovaryum‘un 1/ 3 ile 1/2 ‗si kadar bir
büyüklüğe ulaĢır. Sarımtrak renkteki luteal hücreler dördüncü ayın sonuna
değin
progesteron
salgılamayı
sürdürür.
Bundan
sonra
plasentanın
trofoblastik bileĢeni tarafından salgılanan progesteron hormonunun miktarı
gebeliğin sürdürülmesine yetecek düzeye geldiğinden yavaĢ yavaĢ dejenere
olur. 4. aydan önce gebelik korpus luteum‘unun dejenerasyonu genelde
düĢükle sonuçlanır. 18
23
2.4.OVARYUM FİZYOLOJİSİ
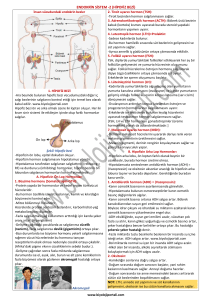
Kadın hormonal sistemi, üç ayrı hormon grubundan oluĢur.
Bunlar:
1) Daha önceleri luteinizan serbestleĢtirici hormon olarak da
isimlendirilen,
bir
hipotalamik
serbestleĢtirici
hormon,
Gonodotropin SerbestleĢtirici Hormon (GnRH)
2) Ön hipofiz hormonlarından Follikül-Uyaran Hormon (FSH)
ve Luteinize edici Hormon (LH). Bu hormonların her ikisi de
Hipotalamusta sentezlenen GnRH – serbestleĢtirici hormona
yanıt olarak salgılanırlar.
3) Ovaryum hormonları; östrojen ve progestron. Bu hormonlar
ön hipofiz bezinden salgılanan iki hormona (FSH, LH) yanıt
olarak ovaryum tarafından salgılanırlar.
Kadında menstrual döngü sürecinde, bu hormonların salgılanma
miktarları değiĢiklik gösterir. Döngünün farklı evrelerinde son derece farklı
hızlarda salgılanırlar. Hipotalamustan GnRH salgısı artıĢ ve azalıĢ Ģeklinde
büyük değiĢiklikler gösterirken, bu artıĢ ve azalıĢlar aylık döngü sürecinde
daha az ya da çok küçük düzeylerde bulunmaktadır. Hormonların ortalama
her 1-3 saatlik kısa devrelerde salgılandığı bilinmektedir.
29,30
Menstrual döngü sürecinde ovaryumdaki değiĢikliklerin tümü
hipofiz bezi ön lobundan salgılanan gonadotropik hormonlar; FSH ve LH ‗a
bağlıdır. Gonadotropik hormonlarla uyarılmayan ovaryumlar inaktiftir. FSH ve
24
LH molekül ağırlıkları yaklaĢık 30.000 olan küçük glikoprotein yapıda
hormonlardır. 30
FSH ve LH, ovaryumlardaki hedef hücrelerini, hücre zarlarında
bulunan son derece özgül reseptörlere bağlanarak uyarırlar. Aktive olan
reseptörler bu hücrelerin salgılama, büyüme ve çoğalma hızını artırırlar. Bu
uyarıcı etkilerin hemen tümü hücre sitoplazmasındaki ikincil haberci siklik
AMP( cAMP) sisteminin aktivasyonu sonucu gerçekleĢir. cAMP protein kinaz
oluĢumunu sağlar daha sonra da çeĢitli fosforilasyonlarda anahtar enzim
görevi yaparak pek çok hücre içi fonksiyonu hızlandırır.
30
Ovulasyon öncesinde, folliküllerden birisi daha fazla büyümeye
baĢlar. Bu follikülde bol miktarda östrojen salgılanır ve östrojen follikül
üzerinde pozitif geribildirim etkisi yapar. FSH, granüloza ve teka hücrelerinin
çoğalmasını artırır. Bunun sonucunda daha fazla östrojen salgılanır ve
yeniden hücresel proliferasyon döngüsüne girilir. FSH ve östrojenlerin
birleĢimi granüloza hücrelerinde FSH ve LH reseptör sayısının artmasına
neden olur. Ayrıca teka hücrelerinde daha az bir pozitif geribildirim etkisi
yaratır. Bu etkiler birlikte, folliküler sıvı ve hormon salgılanmasının hızla
yükselmesine neden olur. Bunun yanında, follikülden salgılanan büyük
miktarlardaki
östrojen
hipofiz
bezi
ön
lobundan
daha
fazla
FSH
salgılanmasını baskılamak için hipotalamusu etkiler. Bu yolla kendi pozitif
geribildirim uyarısıyla geride kalan az geliĢmiĢ folliküllerin daha da geliĢmesini
engellediği düĢünülmektedir. Bu nedenle en büyük follikül pozitif geribildirim
etkiyle büyümesini sürdürürken, diğer folliküller büyümeyi durdurur ve geriler.
29,30
25
LH follikül büyümesinin son evresinde ve ovulasyon anında
gerekli bir hormondur. Bu hormon olmadığında çok fazla miktarlarda FSH
olsa bile, follikül ovulasyon evresine kadar geliĢemez. Ovulasyondan yaklaĢık
2 gün önce hipofiz ön lobunun LH salgı hızı belirgin bir Ģekilde yükselir. Bu
artıĢ ovulasyondan yaklaĢık 16 saat önce 6 ile 10 katı kadardır. FSH da aynı
süreç içinde 2-3 kat artar. Böylece iki hormon aynı anda, birlikte etki ederek
ovulasyon öncesi son birkaç gün içinde, follikülün hızla büyümesini sağlar. LH
ayrıca granüloza ve teka hücreleri üzerine de etki ederek bu hücreleri daha
çok progesteron ve daha az östrojen salgılayan hücrelere dönüĢtürür. Bu
nedenle ovulasyondan yaklaĢık bir gün önce östrojen salgısı azalırken bir
miktar progesteron salgılanmaya baĢlar. 29,30
Ovulasyonu baĢlatan neden hipofiz bezi ön lobundan aĢırı
miktarda LH salgılanmasıdır. LH öncelikle az miktarda progesteron içeren
follikülden folliküler steroid hormonların hızla salgılanmasına neden olur.
Birkaç saat içinde ovulasyon için gerekli iki önemli olay geliĢir: 1- Teka
hücrelerinin lizozomlarından proteolitik enzimler salgılanmaya baĢlar. Bu
enzimler kapsül duvarının çözülmesine ve duvarın zayıflamasına neden olur.
Böylece tüm follikül daha fazla ĢiĢer ve stigma dejenere olur. 2- Aynı anda
follikül duvarında hızla yeni kan damarları oluĢurken folliküler dokuda
prostaglandinler (vazodilatasyon oluĢturan yerel hormonlar) salgılanır. Bu iki
etki follikülün ĢiĢmesinde katkısı bulunan plazma sızıntısına yol açar.
Sonunda follikülün ĢiĢmesi ve eĢ zamanlı olarak stigmanın dejenerasyonu
follikülün yırtılmasına ve oositin dıĢarı atılmasına neden olur.
29,30
Oositin follikülden atılmasını izleyen ilk birkaç saat içinde geride
kalan granüloza ve teka interna hücreleri hızla lutein hücrelerine dönüĢür.
26
Hücrelerin çapları iki katı ya da daha fazla artar, içlerine dolan lipid
inklüzyonları nedeniyle sarımsı bir renk alır. Bu sürece luteinizasyon, oluĢan
hücre kütlesine de korpus luteum adı verilir. 29,30
Korpus luteum granüloza hücrelerinin sitoplazmaları granülsüz
endoplazmik retikulumdan zengindir. Bunlar çok miktarda progesteron ve
östrojen oluĢtururlar. Teka hücreleri ise kadın seks hormonlarından çok,
androstenedion ve testesteron gibi belirli androjenleri sentezlerler. Ancak,
bunların çoğunluğu granüloza hücrelerinde kadın hormonlarına dönüĢtürülür.
30
Granüloza ve teka interna hücrelerinin lutein hücrelerine
dönüĢümü baĢlıca hipofiz bezi ön lobundan salgılanan LH‘a bağımlıdır. LH‘a
―sarımsı renk verme özelliği‖ nedeniyle luteinleĢtirici adı verilmiĢtir. Granüloza
hücrelerinin luteinizasyonu aynı zamanda oositin follikülden atılmasına da
bağımlıdır. Luteinizasyon olayını ovulasyona kadar denetleyen bir diğer
etken, follikül sıvısında bulunan luteinizasyonu baskılayıcı faktördür. Bu
nedenle ovulasyon olaylanıncaya değin follikül içinde korpus luteum
geliĢmez. 30
Özellikle ovulasyon evresinde salgılanan LH öncelikle granüloza
ve teka hücrelerinde luteinizasyona neden olur. Yeni oluĢan lutein hücreleri
sırasıyla çoğalma, geniĢleme, salgı ve dejenerasyona karĢın programlanır.
Hipofiz bezi ön lobundan LH salgılanmasa bile bu süreç devam eder. Olay
ancak 4 ile 8 gün sonra sonlanır. Diğer yandan LH varlığında korpus
luteumun büyümesi artar, salgılanması çoğalır. LH ile hemen hemen aynı
27
özelliklere sahip bir diğer hormon koryonik gonadotropindir. Plasentadan
salgılanan bu hormon korpus luteum‘a etki ederek yaĢamını uzatır ve
genellikle gebeliğin 2-4 aylık süresinde devamını sağlar. 30
Menstrual döngünün luteal evresinde korpus luteumdan salınan
özellikle östrojen ve daha az miktarda progesteron hipofiz bezi ön lobu
üzerinde önemli geribildirim etki yaratarak, FSH ve LH‘nun dozda
salgılanmasını sürdürür. Ayrıca luteal hücreler az miktarlarda da olsa inhibin
hormonu salgılarlar. Bu hormon hipofiz bezi ön lobundan özellikle FSH
salgısını baskılar. FSH ve LH‘nun kanda düĢük düzeylere inmesi ve bu
hormonların kaybolması halinde korpus luteum tamamen dejenere olur. Bu
olaya korpus luteum‘un involusyonu denir. Korpus luteum‘dan östrojen,
progesteron ve inhibin salgılanmadığı anda hipofiz bezi ön lobu üzerindeki
baskılayıcı geribildirim etkisi ortadan kalkar ve bez yine bir miktar FSH ve
birkaç gün sonra da yavaĢ artıĢlarla LH salgılamaya baĢlar. FSH ve LH yeni
bir menstrual döngüde yeni folliküllerin büyümesini baĢlatır. Ancak bu
folliküller yeterince geliĢme göstermezler. Östrojen ve progesteron yokluğu
uterusta menstrüasyona yol açar. 30
2.4.1.Ovaryum
Hormonlarının
İşlevleri
–
Östradiol
ve
Progesteron
Ovaryum‘un seks hormonları, östrojenler ve progestinler olarak
iki tiptir. Östrojenlerin en önemlisi östradiyol hormonu, progestinlerin en
önemlisi de progesteron‘dur. Östrojenler baĢlıca vücutta sekonder kadın seks
özelliklerini veren özgül hücrelerin çoğalma ve büyümesini sağlarlar. Diğer
28
yandan progestinlerin tümü uterus‘un gebeliğe, meme bezlerinin de
laktasyona son hazırlıklarını yürütür. 29
2.4.1.1.Östrojenler
Gebe
olmayan
normal
bir
kadında
östrojenler
yüksek
miktarlarda yalnız ovaryum‘lardan, az miktarlarda da böbreküstü bezi
korteksinden salgılanırlar. Gebelikte ise çok fazla miktarlarda plasentadan
salınırlar. 29,30
Kadınlarda plazmada üç tip östrojen görülür: β-östradiyol, östron
ve östriyol. Ovaryumlardan serbestleĢen en önemli östrojen β-östradiyol‘dür.
Bu organların östron salgısı çok az miktardadır. Östron çoğunlukla,
böbreküstü bezi korteksi ve ovaryum teka hücrelerinden salgılanan
androjenlerin periferik dokulardaki dönüĢümünden kaynaklanır. Östriyol zayıf
bir östrojendir ve özellikle karaciğerde östradiyol ve östron‘un oksidasyon
ürünü olarak ortaya çıkar. 29,30
β-östradiyol‘ün östrojenik erki östron‘a karĢın 12 kat, östriyol‘e
ise 80 kat daha fazladır. Bu nedenle Β-östradiyol‘ün en önemli östojen
olduğu, buna karĢın östron‘un östrojenik etkisinin ihmal edilebilir düzeyde
olduğu söylenebilir. 30
29
Karaciğer, östrojenleri glukuronid ve sülfatlar halinde bağlar.
Bağlı ürünlerin beĢte biri safra ile geri kalanının çoğu idrar ile atılır. Bunun
yanında, karaciğer kuvvetli östrojenler, östradiyol ve östronu tümüyle etkisiz
östrojen olan östriyol‘e dönüĢtürür. Bu nedenle, karaciğer fonksiyonlarının
zayıflamasında, vücutta östrojen erkinin artarak hiperöstrinizm‘e yol açar.
30
2.4.1.2.Östrojenler‘in ĠĢlevleri
Östrojenler çocuklukta çok az miktarda salgılanırlar. Pubertede
ise salgılanan miktar hipofiz gonadotropik hormonların etkisi altında 20 kat ya
da daha yüksek oranda artıĢ gösterir. Bu sırada ovaryumlar, tuba uterina,
uterus ve vajinanın boyutları artar. 30
Östrojenlerin buna ek olarak, vajina epitelini kübik Ģekilden, çok
katlı epitele dönüĢtürerek, puberte öncesine karĢın travma ve enfeksiyonlara
karĢı daha dirençli hale getirir. 30
Puberte sonrası birkaç yıl içinde uterus‘un boyutları iki ya da üç
kat artıĢ gösterir. Daha da önemlisi östrojenlerin etkisiyle endometriyumda
olaylanan
değiĢimlerdir.
Östrojenler
endometriyal
stromada
belirgin
proliferasyona ve endometriyal bezlerde büyük geliĢime yol açarak, daha
sonraki günlerde implante olacak blastosistin beslenmesine yardımcı olurlar.
30
30
Östrojenler, tuba uterinalardaki mukoza katmanlarına da uterus
endometriyumuna benzer etkiler yaparlar. Tuba uterinada silyalı epitel
hücrelerinin sayısında artıĢ sağlarlar. Ayrıca döllenmiĢ oositin uterus‘a doğru
hareketini kolaylaĢtırırlar. 29,30
Östrojenler,
memede
stromal
dokuyu
geliĢtirir,
kanal
sistemlerinin geliĢmesini ve memede yağ birikmesini sağlarlar. Meme
bezlerinde lobul ve alveollerin geliĢimi tek baĢına östrojen etkisi altında
yetersiz düzeydedir. Bu yapıların geliĢimini tamamlaması ve iĢlevsel
özelliklerini kazanması için progesteron ve prolaktinin desteği gereklidir.
Östrojenler
osteoblastik
erkin
artmasına
neden
29,30
olurlar.
Pubertede kadınların üreme çağına girmesiyle birlikte birkaç yıl büyüme çok
hızlıdır. Bunun yanında, östrojenlerin iskelet büyümesi üzerinde bir diğer
etkisi, uzun kemiklerin cismi ile epifizlerin erken birleĢmesine yol açmasıdır.
Sonuç olarak kadında büyüme olayı genelde erkeğin büyümesinden daha
erken yıllarda kesilir. Östrojenden yoksun, önükoid kadınlarda epifizler erken
kapanmadığından, boyları normal eriĢkin kadınlardan daha uzun olur.
30
Östrojenler toplam vücut proteinini hafifçe artırırlar. Östrojen
uygulamasından sonra azot dengesinin pozitif tarafa kayması da bunun
kanıtıdır. Östrojenin seksüel organlar, kemikler ve vücudun daha birçok
dokusunda büyümeyi hızlandırıcı etkisi de bunu göstermektedir.
29,30
31
Östrojenler metabolik hızı hafif de olsa artırırlar. Ancak bu artıĢ
erkek seks hormonu testesteronun etkisine karĢın 1/3 oranda daha düĢüktür.
Bunun yanında hormonun deri altı yağ oranını artırıcı etkisi de vardır.
Östrojenler yağın göğüs ve deri altından baĢka kalçalarda birikmesine yol
açar ki, bu da kadın vücuduna özgü görünümünü kazandırır. 29,30
Östrojen hormonlarının kimyasal yapılarının adrenokortikal ve
suyun geri emilimine neden olduğu belirtilmektedir.
30
Östrojenler salgılandıktan sonra, hedef hücrelere ulaĢmadan
önce birkaç dakikalık süre için dolaĢımda kalırlar. Hücrelere girdikten sonra
10-15 saniye içinde sitoplazmada bir reseptör proteini ile birleĢirler ve
çekirdeğe geçerler. Bu da kromozomal DNA‘nın özgül bölümüyle etkileĢerek
transkripsiyon iĢlemini baĢlatır. Böylece birkaç dakika içinde RNA üretimi
baĢlar. Birkaç saat sonra yeni DNA oluĢabilir ve hücre bölünmesine gidilir.
30
2.4.1.3.Progestinler
Progestinler içinde en önemli hormon progesteron‘dur. Az
miktarlarda bulunan bir diğer progestin 17-α-hidroksiprogesteron‘dur. Bu
hormon progesteronla birlikte salgılanır ve aslında onunla aynı etkiye sahiptir.
Bu nedenle progesteron tek önemli progestin olarak kabul edilmemektedir.
30
32
Gebe olmayan bir kadında progesteron ovaryum döngüsünün
yalnızca ikinci yarısında, korpus luteum‘dan salgılanır. Döngünün ilk yarısında
plazmada bulunan az miktardaki progesteron ise eĢit olarak ovaryumlar ve
adrenal korteks‘den salgılanır. Ancak gebelik sırasında, özellikle gebeliğin
4.ayından sonra plasentadan büyük miktarlarda progesteron serbestleĢir.
Progesteron hormonu
30
salgılandıktan birkaç dakika sonra
tümüyle progesteron etkisi olmayan diğer steroidlere yıkılır. Hormonun
değiĢimi özellikle östrojenler için geçerlidir. Karaciğerin bu metabolik yıkımda
özel bir yeri vardır. Progesteronun en önemli yıkım ürünü pregnandiyol‘dür.
Özgün progesteronun yaklaĢık %10‘u idrarla, bu Ģekliyle atılır.
29,30
2.4.1.4.Progesteronların ĠĢlevleri
Progesteronun en önemli iĢlevi, kadın menstrual döngüsünün
ikinci yarısında uterus endometriyumunda salgılama ile ilgili değiĢimleri
baĢlatarak uterus‘u blastosistin implantasyonuna hazırlamaktır. Uterus
kasılmalarının Ģiddetini azaltarak implante blastosistin atılmasını engeller.
29,30
Progesteron alveol hücrelerinin proliferasyonuyla memelerdeki
lobül ve alveollerin geliĢimini hızlandırır. Böylece göğüsler büyüyerek
salgılayıcı
bir
yapı
kazanırlar.
Progesteron
alveollerin
süt
salgısını
sağlayamaz. Süt ileri dönemlerde memeler hazırlandıktan sonra hipofiz bezi
ön lobundan salgılanan prolaktin hormonunun uyarısıyla salgılanabilir.
29,30
33
Büyük
miktarlardaki
progesteron
hormonu
böbrek
distal
tubuluslarından sodyum, klor ve suyun geri emilimini artırabilir. Tübül epitel
hücrelerinden sodyum iyonlarının taĢınmasını sağlayan progesteron ve
aldosteron aynı reseptör proteinlerine bağlanırlar. Progesteron protein
moleküllerine
bağlandığında
aldosteron
bağlanamaz.
Progesteron
aldosteronun etkisine karĢın çok daha düĢük düzeyde sodyum taĢınımına yol
açar. Bu nedenle, progesteron böbrek tübüllerinden sodyum ve suyun geri
emilimini ancak düĢük düzeyde etkileyebilir. Hormonun etkisi daha çok
aldosteronun etkisini baskılamaktadır. Bu Ģekilde vücutta belirgin bir su ve
sodyum kaybı oluĢur. 30
2.4.2.Hipotalamus’tan Salgılanan GnRH ve Bu Hormonun
FSH ve LH Salgılanmasına Etkileri
Ön hipofiz hormonlarının çoğunluğu, hipotalamusdan salgılanan
―serbestleĢtirici hormonlar‖ tarafından denetlenir. Bu hormonlar hipofiz bezi
ön
lobuna
hipotalamik-hipofizier
portal
sistem
yoluyla
taĢınırlar.
Gonodotropinlerin uyarılmasında bir serbestleĢtirici hormon olan GnRH çok
önemlidir. 30
Deneysel araĢtırmalara karĢın, hipotalamus‘un GnRH salgısı
sürekli değildir. Salgılama her 1-3 saatte bir gerçekleĢir ve birkaç dakika
süreyle devam eder. 30
34
GnRH sürekli uygulandığında ön hipofiz bezinin FSH ve LH
salgısı kaybolur. Buna bağlı olarak GnRH aralıklı salgılanmasındaki özellik
bilinmeyen nedenlerle hormonun iĢlevsel olması açısından önemlidir.
GnRH‘nın aralıklı salgısı, aynı Ģekilde aralıklı LH atılımına neden olur.
30
GnRH‘nın hipotalamusdan aralıklı salgılanması aynı Ģekilde
FSH salgısının azalmasına ya da artmasına neden olur. Buradaki önemli
nokta, GnRH‘nın FSH üzerine etkisinin uzun süreli olmasıdır.
30
35
2.5.İNFERTİLİTE VE OVULASYON İNDÜKSİYONU
2.5.1.İnfertilite
Herhangi bir doğum kontrol yöntemi uygulamadan, haftada en
az iki kez düzenli bir cinsel yaĢama rağmen, bir yıl süreyle gebelik
oluĢmamasına infertilite denir. Ġnfertilite sorununun üreme çağındaki çiftlerin
yaklaĢık %10-15‘ ini etkileyen bir faktör olduğu bilinmektedir. Ġnfertilite primer
ve
sekonder
olarak
iki
Ģekilde
değerlendirilir.
Daha
önce
gebelik
oluĢmamasına primer infertilite, ancak daha önce gebelik oluĢmasına karĢın
bir baĢka gebeliğin oluĢmamasına sekonder infertilite adı verilir. Kadın
yaĢamının menapoz, puberte, gebelik gibi evrelerinde gebelik oluĢması
zordur ya da olası değildir, buna da fizyolojik infertilite adı verilir.
31,32
Ġnfertilitenin en sık görülen nedenleri; ovulatuvar bozukluk
(%15), tubal ve peritoneal patoloji (%30-40) ve erkek faktörleridir (%30-40).
Geri kalanı ise nedeni açıklanamamıĢ infertilitedir. Her bir olgunun sıklığı
yaĢla değiĢmektedir. Genç kadınlarda ovulasyon bozuklukları daha sık
görülürken, tubal ve peritoneal faktörler genç ve yaĢlılarda eĢit sıklıktadır.
Erkek faktörleri ve nedeni açıklanamayan infertilite ise ileri yaĢlardaki çiftlerde
daha sık görülmektedir. 33
Döllenmenin oluĢması için oosite ulaĢabilecek ve oositi
dölleyebilecek normal, hareketli ve yeterli sayıda spermiyum olması, eĢ
zamanlı uygun oosit salınımı, spermin oosite ulaĢabilmesi ve zigot ya da
36
embriyonun uterus‘a ulaĢabilmesi için serbest bir pasajın olması ve
implantasyona
izin
verecek
olgun
bir
endometriyumun
bulunması
gerekmektedir. Bu evrelerdeki herhangi bir defekt gebelik oluĢumunu
engelleyecektir. 6
Kadınlardaki en önemli infertilite nedeni yaklaĢık olarak %40
oranında ovulasyon yetersizliğidir. Ovulasyon yetersizliği anovulasyon veya
ciddi oligoovulasyon Ģeklinde olabilir. Dünya Sağlık Örgütü (WHO) ovulatuvar
bozuklukları üç ana gruba ayırmıĢtır:
Grup I: Hipotalamik hipofizer yetmezlik
DüĢük gonadotropin ve östrojen hormon düzeyleriyle seyreden
hipogonadotropik hipogonadizmi, normal prolaktin yoğunlukları olan ve
―progesteron challenge‖ testine yetersiz kanama ile yanıt veren hastalardır.
Bu grup stres, kilo kaybı, egzersiz, hipotalamik amenore, strese bağlı
amenore, anoreksia nevroza, Kallmann sendromu ve izole gonadotropin
eksikliği olan hastalardır. Bu hastalarda hipogonadotropik hipogonadizm
bulunmaktadır. FSH ve östrojen düzeyleri düĢüktür, prolaktin normaldir.
Grup II: Hipotalamik hipofizer disfonksiyon
Normogonadotropik,
normoöstrojenik,
anovulatuar,
oligomenoreik hastaları kapsamaktadır. Polikistik over sendromu (PKOS) bu
gruba girmektedir. Bu gruptakiler tipik olarak normal FSH, östrojen ve
prolaktin yoğunluğuna sahiptirler.
37
Grup III: Ovaryan yetmezlik
Bu
gruptaki
hastalarda
hipergonadotropik
hipogonadizmle
birlikte düĢük östrojen düzeyleri vardır. Ovaryan yetmezlik ve ovaryan
rezistansların tüm türleri bu gruba girmektedir. Yüksek FSH yoğunluğuna
sahiptirler. 6,34
1960‘lı yılların baĢlarına değin anovulasyon ve diğer infertilite
tiplerinin etkili tedavisi tam olarak bilinmiyordu. 1970‘li yıllarda anovulasyon
tedavisi için gonadotropin salgılatıcı hormon (GnRH) ve prolaktin baskılayıcı
ajanlar olarak iki seçenek öne sürülmüĢtür. GnRH izole edilmiĢ ve yapısı
tanımlanmıĢtır. Kısa bir süre sonra laboratuvarda doğal GnRH preparat
sentezi ile klinik kullanım için daha uygun hale gelmiĢtir. Hipofiz GnRH
tedavisinin hipotalamik kökenli hipogonadotropik amenoreli kadınlarda
ovulasyon indüksiyonu ve gebelik gerçekleĢmesinde çok etkili olduğu
kanıtlanmıĢtır. 15,34,35,36,37,38,39
2.5.2.Ovulasyon İndüksiyonu
Günümüzde ovulasyon indüksiyonunda amaç
anovulatuar
hastalarda ovulasyonu baĢlatmaktır. Ovulasyon sağlanabildiği koĢullarda ise
esas amaç tek folliküllü ovulasyondur. Günümüzde 3 değiĢik ovulasyon
indüksiyonu uygulanmaktadır:
1) Yerine Koyma (Subtitution) Tedavisi: Gonadotropin eksikliği
olan amenoreik kadınlara (WHO grup I) uygulanan tedavidir.
2)
Düzenleme
tedavisi:
Oligoovulasyonu
veya
persiste
anovulasyonu olan kadınlara uygulanan tedavidir. (PKOS gibi).
38
3) Denetimli Ovaryan Hiperstimülasyon (KOH): Bu yöntem ise
çoklu
infertilitesi
(hafif
endometriozis,
geçici
anovulasyon,
hafif
oligoastenospermi, tek taraflı tubal oklüzyon gibi nedenlerin kombinasyonu)
olan ya da nedeni açıklanamayan infertilitesi bulunan hastalara uygulanmakta
olan tedavidir ve herzaman intrauterin inseminasyon ve invitro fertilizasyon ile
birlikte kullanılır. 15
2.5.2.1.Gonadotropinler İle Ovulasyon İndüksiyonu
Ovulasyon indüksiyonu için kullanılan gonadotropinler 3 türdür.
Bunlar; üriner, saf üriner ve rekombinant. YaklaĢık 30 yıl süresince tek
gonadotropin
tipi
insan
menapozal
gonadotropin‘i
olmuĢtur
(hMG,
menotropin). Yeni hMG preparatları eskilerinden daha iyi saflaĢtırılmıĢtır ve
cilt altı uygulanabilirler. 20 yıl önce anti-hCG antikorları ile yapılan
immünoaffinite yöntemi sayesinde üriner LH uzaklaĢtırılarak daha iyi
saflaĢtırılmıĢ üriner FSH elde edilmiĢtir. 40
FSH ve LH‘nın granuloza ve teka hücreleri üzerindeki etkileri
çok iyi nitelendirilmiĢtir ve folliküler geliĢim için esansiyeldir. Etki mekanizması
klomifen sitrat‘a karĢın daha iyi bilinmektedir. Tedavinin baĢarılı olabilmesi
için ovaryumların iĢlevsel olması gerekir, çünkü verilen HMG endojen
gonadotropinlerin
yerini
alarak
ovulasyon
indüksiyonu
yapmaktadır.
Kendiliğinden olan döngülerde olduğu gibi FSH birincil olarak granüloza
hücreleri ve LH teka lutein hücreleri üzerinde etki göstererek folikülogenezisi
uyarırlar. 41
39
Ġnsan menapozal gonadotropin‘i (HMG) üriner bir üründür ve
postmenopozal kadınların idrarlarından elde edilmektedir. EĢit oranda FSH ve
LH içermektedir. LH‘nın yüksek düzeyde olması ve bu nedenle fertilizasyon
ve implantasyonun negatif olarak etkilenmesi FSH içeriği baskın olan yeni
gonadotropin ürünlerinin (uFSH) geliĢmesine yol açmıĢtır. uFSH‘da üriner bir
üründür ve idrar proteinleri ile kontaminedir.
41
Günümüzde gonadotropin preparatları artık 0.001 IU ‗dan az LH
içermektedirler ve üriner protein içerikleri de cilt altı uygulanabilir. YaklaĢık 10
yıl önce genetik mühendisliği sayesinde in vitro rekombinant FSH ( rFSH)
üretilmiĢtir. rFSH insan FSH gen kodları ile transfekte edilmiĢ hamster
ovaryum
hücrelerinden
üretilmektedir.
Rekombinant
gonadotropinlerin
avantajı daha az üriner protein içermeleri, daha tutarlı bir yapılarının olması
ve biyolojik erklerinin kiĢiden kiĢiye çok daha az değiĢiklik göstermesidir.
40,42,43
Ġnsan koryonik gonadotropin (hCG) ise, karbonhidrat yan zincirli
glikoprotein yapılı bir hormondur. LH, FSH ve TSH‘nın heterodimerik yapısı
ile benzerlik gösterir. Bu hormonların α-altbirimleri ortaktır, β-altbirimleri ile
ayrılırlar. hCG 25 yıldan beri folliküler olgunlaĢmanın sonlandırılmasında
kullanılmaktadır. Mazer ve Ravetz gebe kısrakların serum gonodotropinlerini
ve koyunların hipofiz ekstrelerini follikül uyarımı için ve hCG‘yi folikül
olgunlaĢmasının sonlandırılması için amenoreik kadınlarda kullanmıĢlar ve
hipofizer bir faktörün folikül geliĢimini uyardığı ve hCG‘nin ovulasyonu
indüklediği sonucuna varmıĢlardır. 42,44,45
40
2.5.2.1.1.Gonadotropinlerin Endikasyonları
Hipogonadotropik hipogonadizmi olan bireyler (hipotalamik
amenore, WHO Grup I) ekzojen gonadotropin ile en çok tedaviye aday grubu
oluĢtururlar. Klomifen sitrat ve benzer tedavi yöntemleri hipotalamik-hipofizerovaryum
aks
düzgün
olmadığında
etkisizdir.
Hipogonadotropik
hipogonadizmli bir kadında ilaç seçimi insan Menapozal Gonadotropin (hMG)
olmalıdır çünkü hem FSH hem de LH içermektedir.
46
Klomifen sitrat‘a dirençli anovulasyon durumunda; Klomifen
sitrat ile gebelik sağlanamadığında ekzojen gonadotropin tedavisi açık bir
seçenektir. Klomifen sitrat‘a dirençli PKOS‘lu( WHO Grup II) hastalarda serum
gonadotropinleri normaldir. Çoğunlukla LH düzeyi de yüksektir. Bu hasta
populasyonunda tedavi ile ekzojen gonadotropin, endojen gonadotropinlerle
üst üste gelir. Açıklanamayan Ġnfertilite olgularında ekzojen gonadotropinler
ile tedavide amaçlanan birden fazla olgun oositin sağlanmasıdır.
47
2.5.2.1.2.Gonadotropinlerle Tedavi Rejimi
Hipogonadotropik hipogonadizmli (WHO grup I) ve klomifen
sitrat‘a dirençli (WHO Grup II) hastalarda düĢük doz ile indüksiyona (75 IU/
gün) step-up protokolü çerçevesinde baĢlanabilir. 4-7 günlük bir uyarma
süresi sonunda serum östradiol düzeyine ilk yanıtı ölçmek için bakılmalıdır.
Daha sonra gonadotropin dozu aynen sürebilir ya da artırılabilir. Serum
östrodiol düzeyi bir kez yükseldikten sonra, geliĢen folliküllerin sayısını ve
çapını ölçmek için 1-2 günde bir transvajinal ultrasonografi yapılır. Önde
41
giden follikül çapı 15-18 mm‘ye ulaĢtığında ovulasyonu sağlamak için hCG
enjeksiyonu yapılır. Genelde 36-48 saat sonra ovulasyon beklenir. 47
PKOS‗lu kadınlar çoğu zaman gonadotropinlere aĢırı duyarlılık
gösterebilirler. Bu kadınlarda genellikle FSH uyarısına yanıt verecek çok
sayıda antral follikül vardır. 47
Ovaryan hiperstimulasyon sendromu (OHSS) ve çoğul gebelik
riski daha düĢük dozları içeren ( 37.5- 75 IU / gün) ‗düĢük-yavaĢ‘ (low dose
step-up) tedavisi ile engellenebilir. 48
Alternatif olan step-down rejiminde ise tedavi yüksek dozda
baĢlar ( 150-225 IU/gün) ve seçilmiĢ folliküller içindeki duyarlılığı düĢük
folliküller gerilerken, duyarlılığı yüksek folliküllerin büyümeye devam etmesi
için giderek dozların düĢürülmesi esasına dayanır. 49
Klomifen sitrat‘a dirençli ve açıklanamayan infertilite hastalar için
ardıĢık klomifen sitrat ve gonadotropin tedavisi uygun görülmektedir. Tipik
döngüde standart klomifen sitrat tedavisini izleyerek (50-100 mg/gün)
klomifen sitrat tedavisinin son günü düĢük doz gonadotropin tedavisi (75 IU
FSH veya hMG) uygulanır. Daha sonra sıradan bir gonadotropin ile
indüksiyon tedavisinde olduğu gibi izlenir. 49
42
2.5.2.1.3.Gonadotropin Tedavi Sonuçları
Bir menstruel döngü içerisinde gebelik elde edebilme olasılığı
artan östradiol ile birlikte artar. Bu durum ovarian hiperstimulasyonu ve çoğul
gebelik olasılığını da artırır. Gonadotropin rejimleri ile en iyi sonuç en yüksek
östradiol yoğunlukları 500-1500 pg/ml arasında olduğunda alınır. Ovaryan
ultrasonografi ile folliküllerin sayısı ve çapı belirlenir. Kendiliğinden olan bir
döngüde, o döngüde rol alacak antral folliküller menstruasyonun 5-7 gün
arası izlenebilir. Baskın follikül 8-12 gün arası belirir ve günde yaklaĢık 1-3
mm büyür, ortalama 20-24 mm iken ovulasyon olur. Kendiliğinden olan
döngülerin %5-10 kadarında iki preovulatuvar follikül geliĢir. 50,51
Ekzojen gonadotropin ile uyarılmıĢ döngülerde baskın follikül
benzer bir büyüme gösterir ancak daha küçük çaplarda olgunluğa ulaĢır.
Follikül çapı arttıkça ovulasyon olasılığı da artar. hCG uygulandıktan sonra,
14 mm ve daha küçük folliküller ender ovulasyon gösterir. Folliküller yaklaĢık
%40‘ı 12-16 mm, %70‘i 17-18mm, %80‘i 19-20mm çapındadır ve aslında tüm
büyük folliküller ovulasyon gösterir. 51
2.5.2.2.Klomifen Sitrat (KS) İle Ovulasyon İndüksiyonu
Klomifen sitrat ilk olarak 1956 yılında sentezlenmiĢtir. Klinik
denemeleri ilk olarak 1960‘ da baĢlatılmıĢtır ve ABD‘ de klinik kullanımı 1967
yılında onaylanmıĢtır. 51
43
Kimyasal olarak klomifen sitrat bir nonsteroidal trifeniletilen
yapısında olup, östrojen agonisti ve antagonisti etkileri birlikte barındırır.
Klomifen sitrat iki farklı steroizomerin karıĢımıdır, farmakolojik olarak cis ve
trans yapıları vardır; enklomifen (cis-klomifen) ve zuklomifen (trans-klomifen)
olarak adlandırılırlar. Enklomifen en etkili izomerdir ve ovulasyonu sağlayan
etkiden sorumludur. Enklomifenin yarı ömrü kısadır ve serum konsantrasyonu
tedaviden sonra hızlıca yükselir ve birkaç gün sonra düĢer. Zuklomifen daha
yavaĢ atılır. Tek dozdan sonra bile haftalarca serumda saptanabilir. ArdıĢık
tedavilerde ovaryumda birikebilir. 51,52,53,54
Klomifen sitrat etkisini östrojen reseptörleri bloke ederek
gösterir. Yapısal benzerliği nedeniyle tüm vücuttaki östrojen reseptörlerine
bağlanır. Ancak östrojenden farklı olarak reseptörlere uzun süre bağlı kalır ve
reseptör dönüĢümünü düĢürerek ortamdaki reseptör sayısını azaltır. Böylece
çekirdekteki östrojen reseptörleri düzeyinde endojen östrojenlerle yarıĢarak
östrojen etkisini engeller. Bu engelleme hipotalamus-hipofiz sisteminde
olduğunda östrojenin negatif geribildirim etkisi ortadan kalkacağından
hipofizden FSH ve LH salgılanması artar. 51,53
2.5.2.2.1.Klomifen Sitrat Endikasyonları
Klomifen sitrat tedavisi anovulatuvar infertilitesi olan kadınlarda
kullanılan ilk sıradaki ilaç seçimidir. Anovulatuvar infertilitede KS tedavisi
sadece dolaĢan östrojen seviyesi gonadotropinler üzerinde negatif geribildirim
oluĢturacak düzeye geldiğinde etkilidir. Progesteron uygulanmasından sonra
oluĢan kanama, dolaĢımdaki yeterli östrojenin klinik yansıması olarak kabul
44
edilebilir. Klomifen sitrat tedavisi genellikle anovulatuvar disfonksiyonlu
kadınlar ile sınırlandırılmıĢ olsa da, luteal evre hasarı ve açıklanamayan
infertilitesi olan, infertilite süresi kısa olan hastalarda da kullanılabilir.
Ovulatuvar infertilitede ise KS multifolliküler geliĢimi uyararak olası bir
ovulasyon disfonksiyonunu düzelterek gebeliği sağlamaktadır.
55,56
Klomifen sitrat‘ın hipotalamus-hipofiz aksı hasarlı kadınlarda ya
da dolaĢan östrojen düzeyi çok az olan hastalarda (WHO Tip I ve III)
kullanılması uygun değildir. 53
2.5.2.2.2.Klomifen Sitrat Dozu
Klomifen sitrat oral olarak uygulanır. Menstrüel döngünün 3 ile
5. günleri arasında uygulamaya baĢlanabilir. Ġlk döngüde tedaviye 50mg/gün
baĢlanır, tedavi 5 gün olarak belirlenmiĢtir. Ovulasyon olmaması durumunda
izleyen döngülerde ilacın dozu 50 mg/gün arttırılır. 150mg/gün ile ovulasyon
elde edilemezse farklı bir ilaca geçilmelidir. Ovulasyon sağlandıktan sonra,
aynı doz gebelik oluĢana değin ya da toplam tedavi süresi 6 döngüyü bulana
değin tekrarlanır. 53,57
2.5.2.2.3.Klomifen Sitrat Tedavi Sonuçları
KS ile yapılan ovulasyon indüksiyonunda %60-90 ovulasyon
görülmüĢtür. Üç baĢarılı ovulasyondan sonraki kümülatif gebelik oranı
45
%70‘dir. Amenoreik kadınlar oligomenoreik kadınlardan daha çok gebe
kalırlar. Bunun nedeni olasılıkla bu hastalarda altta yatan diğer infertilite
nedeni olması ve aslında ovulasyon yapabilmeleridir. Tüm çalıĢmalarda
ovulasyon sağlanmıĢ tedavilerdeki döngü baĢına gebelik oranı %15‘dir. Artan
yaĢa koĢut biyolojik fertilite azaldığından 35 yaĢ üzerinde klomifen sitrat ile
tedavi uygun bir yöntem değildir. 3-6 ovulatuvar döngüye karĢın gebelik
oluĢmaz ise diğer infertilite nedenlerinin gözden geçirilmesi gereklidir. 58,59
2.5.2.2.4.Klomifen Sitrat’ın Yan Etkileri
Birçok çalıĢmada çeĢitli modellerde, KS‘nin endometriyal
büyüme ve olgunlaĢmaya, servikal mukusun kalitesine olan yan etkileri
gösterilmiĢtir. Genel olarak bu etkilerin uzun süreli tedavi ya da yüksek dozda
tedavi sonrası olduğuna inanılır. 60
Klomifen sitrat tedavisinin küçük yan etkileri genelde sıktır ancak
tedaviyi kesmeyi gerektirecek kadar Ģiddetli olması çok enderdir. Kısa dönem
sıcak basmaları izlenmektedir ancak, klomifen sitrat tedavisinin sadece
merkezi olarak endojen östrojen düzeylerinin düĢük olarak algılanmasına
neden olmasından dolayı bu etki geçicidir. Ruh halinde dalgalanmalar sıktır.
Hafif ve seyrek yan etkiler ise meme hassasiyeti, pelvik basınç ya da ağrı ve
bulantıdır. Görme bozuklukları, ıĢığa duyarlılık sık rastlanmayan (%2) ve geri
dönüĢümlüdür. 60
46
2.6.Proliferasyon Hücre Çekirdek Antijeni (PCNA)
PCNA 36 kDa ağırlığında türler arasında iyi korunmuĢ, hücre
döngüsü için esas olan düzenleyicidir. DNA sentezi sırasında DNA
hasarlarının onarılması ve S fazında DNA polimeraz delta için ko-faktör
olarak iĢlem gördüğü gösterilmiĢtir. 61,62,63
Hücre döngüsünün G1 evresinde ekspresyonu baĢlar, en
yüksek düzeye S fazında ulaĢır ve G2/M fazında da azalır. Dinlenme
evresindeki hücreler farklı mitojenlerle uyarıldığında aynı derecede PCNA ve
DNA sentezi yaptığı gösterilmiĢtir. Biyolojik yarı ömrü yaklaĢık 20 saattir.
Aldehitleri kullanarak fikse edilmiĢ hücrelerde ise PCNA‘nın dağılımı farklıdır
ve hücre döngüsü boyunca yoğun diffüz nükleer boyanma saptanır.
61,62,64,65,66,67
PCNA‘ni
hücrelerde
ve
dokularda
göstermeye
yönelik
çalıĢmalara otoimmün hasta serumundan elde edilen poliklonal antikorların
kullanılmasıyla baĢlanmıĢtır. Daha sonra 1990‘da, bakterilerde PCNA‘ni
ortaya çıkararak denatürasyona neden olmayan koĢullarda saflaĢtırılmıĢ ve
bununla farelerin immünizasyonundan elde edilen, 11 yeni monoklonal
antikor tanımlanmıĢtır. Normal eriĢkin dokularında çekirdekte yaygın ve / veya
granüler boyanma Ģeklinde izlenen PCNA hücre proliferasyonu ile birliktelik
göstermiĢtir. 68,69,70
47
PCNA‘nin normal dokularda gösterilmesi proliferatif potansiyeli
yakından
yansıtırken,
neoplazmlarda
daha
karmaĢık
ifade
biçimleri
görülmektedir. Proliferasyonla birlikte olan çeĢitli belirleyicilerin prognozla
iliĢkilerinin farklı tümör tiplerinde değiĢkenlik göstermesi PCNA için de
geçerlidir.
PCNA‘nin
proliferasyonu
immünohistokimyasal
belirleyen
tekniklerin
tümü
yöntemle
prognozla
gösterilmesi
iyi
gibi
korelasyon
göstermektedir. 71,72,73,74
PCNA ovaryumda mayotik bölünme halinde olan hücrelerde,
çekirdek ve
sitoplazmada
saptanmıĢtır. Gonadotropinler ve
östradiol
granüloza hücrelerinin çoğalma evresini baĢlatabilirler. Fare ve sıçanlar ile
yapılan çalıĢmalarda, granüloza hücre çoğalmasının hızlı evresinin antrum
oluĢumu sırasında gerçekleĢtiği gösterilmiĢtir. 63
Yapılan
immünohistokimyasal
çalıĢmalarda;
PCNA‘nin
primordiyal folliküllerde zayıf olmakla birlikte oositte, geliĢen folliküllerde ise
granüloza hücrelerinde ve oositte izlenmiĢtir. 75
Ovaryum‘da in vivo ve in vitro çalıĢmalarda sıçan, sığır ve
domuzlarda PCNA salınımının baĢlangıç ve erken folliküllerde olaylandığı
gösterilmiĢtir. 63,76
48
2.7.C-FOS
c-fos geninin transkripsiyonunu ilk kez 1984 yılında Mike E.
Greenberg ve Ed B. Ziff büyüme faktörleri ile uyardıkları fibroblastlarda
göstermiĢlerdir. C-fos geninin transkripsiyonu hücreleri etkileyerek büyüme ve
farklılaĢmayı sağlayan çok sayıda etken tarafından hızla ve geçici olarak
sağlanır. C-fos bu nedenle hücrede herhangi bir uyarana karĢı ilk yanıt veren
genlerdendir [erken–ivedi geni, (IEG) ya da erken yanıt geni, (ERG)]. Bu
gruba giren genler hücre büyümesi, çoğalması, farklılaĢması ve hücrenin
programlı ölümü (apoptosis) ile ilgili transkripsiyon faktörleridir.
77,78,79,80
IEG grubundan olan c-fos geni çekirdeksel bir transkripsiyon
faktörüdür ve nükleer üçüncü ulak ya da hücresel iĢlevler yönünden esas
anahtar olarak düĢünülmektedir. Bir diğer tanımlama ile ―çekirdeksel
protoonkojen (nuclear protooncegene)‖ olan c-fos hücre çoğalması ve
farklılaĢması sırasında, gen ekpresyonunun aĢamalı düzenlenmesi ile ilgilidir.
Fos proteinleri c-fos geninin ürünü olan çekirdek proteinleridir. Bunların
hücredeki ekspresyonu ve birikimi herhangi bir etki olmaksızın görülebilir
ancak bu ekspresyon ve birikim çeĢitli uyarılara yanıt olarak hücre
aktivasyonu ile birlikte artar. 81,82,83,84
Fos proteinleri yine bir IEG olan c-jun geninin ürettiği Jun
proteinleri ile birlikte transkripsiyon faktör kompleksi Aktivatör Protein-1 (AP-1)
‘ in bileĢenleri olarak birlikte iĢlev görürler. Fos proteinlerinin ekspresyonu tipik
olarak geçicidir. Ancak bu ailenin bazı üyelerinin ekspresyonundaki
yüksekliğin devamının kronik uyaranlar tarafından engellendiği rapor
edilmiĢtir.85
49
Fos benzeri (Fos-like) proteinler Fra (kronik-Fos akrabası
antijen, chronic-Fos related antigen) olarak adlandırılırlar. 86
AP-1 (Activator Protein-1); Fos proteinleri (c-Fos, Fos B, Fra 1
ve Fra 2), Jun proteinleri (c-Jun, Jun B ve Jun D) ve bazı ATF proteinlerinden
(ATFa, ATF-2 ve ATF-3) oluĢmuĢtur. Fos proteinleri, Jun proteinleri ile stabil
dimerler oluĢturur. Oysa Jun proteinleri homodimerler ya da Fos ve ATF
proteinleri
ile
heterodimerler
oluĢturabilir.
Tüm
AP-1
proteinleri
fosfoproteinlerdir ve hiperfosforilasyon ve defosforilasyon onların DNAbağlanma ve transaktivasyon erklerini etkileyebilir.
AP-1
kompleksin
üyeleri
87,88,89,90,91,92,93,94
değiĢik
gen
ekspresyonlarını
düzenlerler. Bu bileĢenler çoğalma, farklılaĢma, programlı ölüm ve onkojenik
transformasyon gibi çok sayıda hücresel süreçlerle kritik olarak ilgilidir. 95,96,97
c-fos transkripsiyonu çok çeĢitli mitojenler ve farklılaĢmayı
uyaran maddeler tarafından uyarılabilir ve bağımsız olarak yeni protein
sentezi oluĢturur. Bu nedenle bu tür genler protoonkojenler ve bunların ürünü
olan proteinler de onkoproteinler olarak adlandırılırlar. Kanser geliĢiminde Fos
proteinlerinin katkısı olduğu bilinmektedir. Bazı tümör tiplerinde c-fos ve c-jun
düzeyinin arttığı gösterilmiĢ ve bu artıĢın artan hücre çoğalmasının bir kanıtı
olacağı için tümör oluĢmasında önemli bir rol oynadığı ileri sürülmüĢtür.
Ancak protoonkojenler yalnız kötü huylu geliĢim ile birlikte değil, normal
büyüme
ve
farklılaĢmayı
protoonkojenlerin
da
düzenlerler.
uygun
GeliĢimsel
anomalilerin
olmayan
50
aktivasyonuyla
yüksek
düzeyde
onkoprotein
üretmelerine
veya
protoonkojenlerin anormal üretimine yol açan bir mutasyona bağlı
olabileceği ileri sürülmüĢtür. 91, 98,99,100
Tüm AP-1 proteinleri büyüme faktörlerine hücresel yanıtın
bazı yönlerini düzenlerler. c-fos protoonkojeninin büyüme faktörleri
tarafından oluĢturulan uyarıların hücre dıĢı ortamdan hücre çekirdeği içine
iletilmesinde esas rolü oynadığına inanılmaktadır.
101
c-fos gibi IEG‘ lere NGF (sinir büyüme faktörü) aksiyonu için
gereksinim olduğu ve örneğin; hasarlanmıĢ retinal ganglion hücrelerinin
rejenerasyonlarıyla
ilgili
olduğu,
bu
nedenle
nöronların
canlılığını
sürdürmesi ve rejenerasyonunun anlaĢılması için c-fos‘un düzenlediği
biyolojik olaylar zincirinin kesin tanımlanmasına çok gereksinim olduğu
bildirilmektedir. 101,102
c-fos, hücrelerin hücre dıĢı değiĢikliklere vereceği yanıtın
büyüklüğünü saptamada da fonksiyonel bir gösterge olarak kullanılır.
103
Pek çok hücre dıĢı etkenin c-fos transkripsiyonunun hızlı ve
geçici olarak artmasını sağlamasına karĢın, bazı geçici, nöral ve hormonal
uyarıyla bu transkripsiyonun azaltılabileceğini gösteren çalıĢmalar vardır.
104
c-fos geni ve Fos proteinlerinin büyüme faktörleri, mitojenler
ve pek çok hücre dıĢı etkenle ekspresyonunun artması ve bu artıĢın
hücredeki büyüme, çoğalma, farklılaĢma ve programlı ölüm gibi çok
önemli biyolojik süreçlerle ilgisi nedeniyle bu gen ve proteinler özellikle
doğumsal anomalilerin, onkogenik transformasyonun ve normal geliĢimin
51
değerlendirilmesi yönünde yapılacak hücresel ve genetik çalıĢmalarda
önemli bir anahtardır. 105,106
Yapılan çalıĢmalarda c-fos‘un steroid metabolizmalı birçok
enzimin ekspresyonunda düzenleyici rol oynadığı ortaya konulmuĢtur. cfos
immunlokalizasyonu
follikül
teka
hücrelerinde
gözlenirken granüloza hücrelerinde yüksek bulunmuĢtur.
düĢük
düzeyde
107
2.8.Lösemi İnhibitör Faktör (LIF)
Lösemi inhibitör faktör (LIF) 43 kDa moleküler ağırlığa sahip
bir glikoproteindir. ÇeĢitli dokularda biyolojik aktivitesi bulunur. LIF
embriyonik kök hücrelerin, primordiyal germ hücrelerinin, periferal
nöronların, osteoblastların, adipositlerin, hepatositlerin ve endotelyal
hücrelerin büyümesini ve farklılaĢmasını düzenler.
108
LIF fibroblastlar, alloreaktif T-lenfositler, mitojen aktiviteli
dalak hücreleri, Krebs ve Ehrlich asit mayi hücreleri tarafından üretilir.
Hücre aktivasyonundan sonra monositler tarafından da üretilir. LIF‘in
mezenĢimal hücrelerde sentezi, IL-1α, TGF-β,EGF ve bFGF tarafından
uyarılır. 108,109
LIF‘in birbirine eklenmiĢ birkaç izoformu tanımlanmıĢtır. LIFD, LIF-M, LIF-T. Tüm izoformlar 3 değiĢik birinci ekson (D,M,T) ve ortak
ikinci ve üçüncü ekson içerir. 110
52
Ġnsan LIF-D, LIF reseptörü ile sinyallenen bir biyolojik aktif
proteindir. LIF-M, ekstrasellüler matrikste aktif olur. LIF-T, intrasellüler bir
protein olarak bulunur. 110
LIF, miyeloid lösemi hücrelerinin çoğunu engelleyen ve
makrofajlara dönüĢmesini uyaran bir etken olarak izole edilmiĢtir.
110,111
Yapılan çalıĢmalar LIF‘nün ovaryum iĢlevinde önemli bir
düzenleyici olabileceğini düĢündürmektedir. 108
LIF mRNA ve proteini ovaryum hücre kültürlerinde düĢük
düzeyde ancak yapıcı olarak ifade olmaktadır. LIF mRNA konsantrasyonu
IL-1α ve TNF-α tarafından zaman ve konsantrasyon bağımlı Ģekilde
yükselmiĢtir. HCG‘nin folliküler sıvıda LIF ekspresyonunu uyardığı yapılan
çalıĢmalarda gösterilmektedir. Aynı zamanda granuloza-lutein hücreleri ve
ovaryum stromal hücreleri LIF mRNA eksprese eder ve protein üretirler.
LIF‘in bu hücrelerdeki bulunuĢu ovulasyon fizyolojisinde önemli rol
oynayabileceğini düĢündürmektedir. 112,113,114
53
3.GEREÇ VE YÖNTEM
3.1.Deney Hayvanları ve Gruplandırma
Bu çalıĢmada Gazi Üniversitesi Laboratuvar Hayvanları
YetiĢtirme ve Deneysel AraĢtırma Merkezi‘nden sağlanan Wistar Albino
cinsi, 36 adet, 20 haftalık, 200-250 gr ağırlığında diĢi sıçanlar kullanıldı.
Denekler laboratuar ortamında 3 hafta süresince 12 saat
gündüz ve 12 saat gece döngüsünde ve 23±2°C‘ lik denetimli sıcaklıkta
tutuldu. Tüm deneklerin su ve besine serbest eriĢimi sağlandı. Bu süre
boyunca tüm deneklerden vaginal smear örnekleri alınarak 3 döngü
boyunca izlendi ve siklusları düzenli olan sıçanlar çalıĢmaya alınarak, her
birinde 6‘Ģar denek bulunan 6 grup oluĢturuldu. Bu gruplara ovulasyon
indüksiyon modelleri uygulandı.
Grup 1:Ovulasyon indüksiyonu sağlamak için hormon uygulaması yapılan
gruba ait kontrol grubu (n=6)
Grup 2 : Hormon uygulaması yapılan grup (n=6)
Grup 3 : Hormon uygulaması yapılan gruba ait gebe grubu (n=6)
Grup 4 : Klomifen Sitrat uygulaması yapılan gruba ait kontrol grubu (n=6)
Grup 5 : Klomifen sitrat uygulaması yapılan grup (n=6)
Grup 6 : Klomifen sitrat uygulaması yapılan gruba ait gebe grubu (n=6)
Hormon uygulamasının kontrol grubu olarak belirlenen 1.
gruba hormon uygulaması yapılan 2. grupta uygulanan enjeksiyonlarla eĢ
zamanlı olarak aynı dozlarda SF verildi. 2. grup olarak belirlenen hormonal
54
uyarım grubuna ise ovulasyon indüksiyonu için insan menopozal
gonadotropin (hMG) uygulandı. Bu gruba alınacak deneklere 4. Menstural
döngünün
diöstrus
evresindeyken
150
IU/kg
insan
menopozal
gonadotropin (hMG) intramuskuler olarak 17:00‘ de verildi. 4. döngünün
diöstrus evresini izleyen proöstrus evresinde ise 75 IU/kg insan koryonik
gonadotropin (hCG) intramuskuler olarak 17:00‘ de uygulandı. Klomifen
sitrat uygulamasının kontrol grubu olarak belirlenen 3. gruptaki deneklerin
oluĢturulabilmesi için, aynı 2. gruptaki denekler gibi hormonal uyarım
yapıldıktan sonra östrus evresinde olduğu belirlenen diĢi sıçanlar 1 gece
erkek sıçanlarla birlikte bırakıldı. Vaginal plak oluĢumu görülen sıçanlar
gebeliğin 0. gününde kabul edilerek gruba alındı. Bu gruptaki deneklerin
de gebeliğinin 6. gününe gelindiğinde implantasyon aĢamasında oldukları
kabul edildi. 4. gruba aynı stres koĢullarının oluĢturulması amacıyla
diöstrus evresindeki deneklere 2 gün süresince lavage yoluyla sadece 2
ml SF verildi. 5. gruptaki deneklere ovulasyon induksiyonun oluĢturulması
ereğiyle 2 ml SF içerisinde 100 g/kg klomifen sitrat çözüldü. Çözelti 4
günlük östrus evresinin diöstrus evresindeki deneklere lavage yoluyla 2
gün boyunca verildi. 6. gruptaki deneklerin oluĢturulabilmesi için, aynı 5.
gruptaki denekler gibi klomifen sitrat uygulaması yapıldıktan sonra östrus
evresinde olduğu belirlenen diĢi sıçanlar 1 gece erkek sıçanlarla birlikte
bırakıldı. Vaginal plak oluĢumu görülen sıçanlar gebeliğin 0. gününde
kabul edilerek gruba alındı. Denekler gebeliğin 6. gününe gelindiğinde
implantasyon aĢamasında oldukları kabul edildi.
Denekler genel anestezi altındayken karın bölgeleri açılarak
ovaryum‘ları çıkarıldıktan sonra intraperitoneal ketamin (44mg/kg) ve
ksilazin
enjeksiyonu
ile
uyutularak
ötenazi
gerçekleĢtirildi.
Tüm
deneklerden immunohistokimyasal inceleme için alınan ovaryum örnekleri
%10‘ luk nötral formaldehite alınarak, alıĢılmıĢ histolojik yöntemlerden
geçirilerek parafin bloklar hazırlandı.
55
3.2.İmmünohistokimyasal Yöntem
Kontrol ve deney gruplarına ait ovaryum dokusu bloklarından
polilizinli camlara 4 µm kalınlığında kesitler alınarak immünohistokimyasal
boyamaları
yapıldı.
ÇalıĢmada
hücre
proliferasyonunun
izlenmesi
amacıyla PCNA ( PC-10) Ab-1, (Lot: 106P8120) primer antikoru, LIF ( N18) primer antikoru (Cat: Sc- 1336, (Lot: #B-1710), C-fos( K-25) primer
antikoru (Cat: Sc-253, Lot: #F-1110) kullanıldı.
Kesitler 37ºC‘deki etüvde bir gece tutulduktan sonra
deparafinizasyonu kolaylaĢtırmak amacıyla etüv ısısı 57ºC‘ye çıkarılarak 1
saat daha bekletildi. Camlar deparafinizasyonu tamamlamak için 2 kez
15‘er dakika ksilolde bırakıldıktan sonra sırasıyla %100‘lük, %96‘lık ve
%80‘lik alkol serilerinden 10‘ar dakika geçirilerek sudan, 2 kez 5‘er dakika
distile sudan geçirilerek alkolden kurtarıldı.
PCNA immünboyanması yapılacak kesitler doku içerisinde
formaldehitin kapattığı reseptör bölgelerinin açığa çıkarılmasını sağlamak
amacıyla mikrodalga fırında 1 M sitrat tamponuna (pH: 6.0) (Cat: AP9003- 500, Lot: 9003LT13610, Lab Vision, Fremont, USA) etkin bırakıldı.
Oda ısısında 20 dakika soğutulduktan sonra 15 dakika süreyle hidrojen
peroksit (Cat: TA-125-HP, Lot: 125HP14119, Lab Vision, Fremont, USA)
uygulandı ve endojen peroksidaz aktivitesi bloke edildi. LIF ve c-FOS
immünboyanması yapılacak kesitlere ise sitrat aĢaması uygulanmadan
direk hidrojen peroksit aĢaması uygulandı. Daha sonra, camlar 3 kez 3‘er
dakika PBS (Phosphate Buffer Saline) (pH: 7.4) ile yıkandıktan sonra
özgün olmayan bağlanmaların engellenmesi amacıyla 5 dakika, Ultra V
Block, PCNA için (Anti-mouse HRP kit Lot: MHL90115, Thermo Scientific,
56
Fremont, USA) LIF için ( Histostain Plus, Lot: 724944A, Ġnvitrogen,USA),
c-FOS için (sekonder kit, LOT: C0910, Santa Cruz Biotechnology,
California,USA)
uygulandı.
Bloklama
aĢamasının
ardından
kesitler
yıkanmadan PCNA ( PC-10) Ab-1, primer antikoru (Lot: 106P8120), LIF (
N-18) primer antikoru Cat: Sc- 1336, Lot: #B-1710), C-fos( K-25) primer
antikoru (Cat: Sc-253, Lot: #F-1110) primer antikorlarına etkin bırakıldı.
Daha sonra camlar, 3 kez 3‘er dakika PBS ile yıkandıktan sonra PCNA
için
Anti-mouse
HRP
kit
(Lot:
MHL
90115)
Thermo
Scientific
Fremont,USA) LIF için Histostain Plus (Lot: 724944A) Ġnvitrogen,USA) ve
c-FOS için sekonder kit (Lot: C0910) Santa Cruz Biotechnology
California,USA) 20 dakika biyotinli sekonder antikor uygulanarak primer
antikora bağlanması sağlandı. Tekrar PBS ile yıkandıktan sonra dokular
enzimin biyotine bağlanması amacıyla 20 dakika PCNA için Anti-mouse
HRP kit (Lot: MHL 90115) Thermo Scientific Fremont,USA) LIF için
Histostain Plus (Lot: 724944A) Ġnvitrogen,USA) ve c-FOS için sekonder kit
(Lot: C0910) Santa Cruz Biotechnology California,USA)
streptavidin
peroksidaz enzim kompleksine etkin bırakıldı.
Camlar tekrar PBS ile yıkandıktan sonra kromojen olarak
PCNA antikoru için
AEC (3-amino-9-ethylcarbazole) (Lot: ASA100318,
Thermo Scientific, Fremont, USA) uygulanarak gözle görülebilen immün
reaksiyonun açığa çıkması sağlandı.LIF primer antikoru için kromojen
olarak
DAB
(3,3‘iaminobenzidine
Tatrahyrochloride-Plus
kit)
(Lot:
720221A, Ġnvitrogen, USA) c-FOS için yine DAB kromojen boyası( Sc2053 Lot: C09110) Santa Cruz Biotechnology California,USA) kullanıldı.
Zemin boyamasında Mayer‘in Hematoksileni (Cat:TA-125-MH, Lot:
AMH70809, Lab Vision, Fremont, USA) kullanıldı. AEC ile boyanan camlar
Ultramount (Cat: TA-125-UG, Lot: VM13518, Lab Vision, Fremont, USA)
ile kapatıldı. DAB ile boyanan camlar alkol ve ksilol serilerinden geçirilerek
entallan ile kapatıldı. Kesitler Leica DM 4000 (Germany) bilgisayar destekli
57
görüntüleme sisteminde, Leica Q Vin 3 programında değerlendirildi ve
resimlendi.
3.3. İstatistiksel Yöntem
Tüm istatistiksel analizler SPSS paket programı (SPSS Inc.
USA) kullanılarak yapıldı. P< 0.05 anlamlı kabul edildi. Veri setini
oluĢturan değerlerin normal dağılım gösterip göstermediği Shaphiro-Wilk
varyansların homojenliği Levene testi ile kontrol edildi. Verilerin normal
dağılım göstermediği, varyansların homojen olmadığı belirlendi. Buna
uygun olarak iki grup ortalamasının karĢılaĢtırılması için Mann–Whitney U,
ikiden fazla grup ortalamasının karĢılaĢtırılması için Kruskal-Wallis testi
uygulandı.
58
4.BULGULAR
4.1.PCNA BULGULARI:
Hormon
uygulaması
yapılan
gruplara
ait
kontrol
ovaryumlarında geliĢim aĢamalarındaki tüm folliküler, atretik folliküller ve
stroma normal yapıdaydı. Primordiyal folliküllerde oosit çekirdek ve
çekirdekçiğinde, follikül hücre çekirdeklerinde PCNA tutulumu belirgin ve
orta dereceliydi. Atretik folliküllerde, granüloza hücreleri ve teka interna
hücrelerinde
izleniyordu
(Resim
1).
Büyük
büyültmeli
resimlerde
primordiyal, tek ve çok sıralı hücre katmanı içeren folliküllerde oosit,
granüloza ve teka interna hücre çekirdeklerindeki immün tepkime belirgin
olarak gözlemleniyordu. Antrumu belirgin sekonder folliküllerde oositteki
PCNA tutulumunun oldukça azaldığı, granüloza hücre tutulumu ile teka
interna hücre tutulumunun ise oldukça yoğun olduğu ayırt ediliyordu. Bazı
stromal hücrelerde de zayıftan ortaya değiĢen boyanma belirgindi. Atretik
folliküllerde ise immünreaktivitenin oldukça kuvvetli olduğu görüldü (Resim
2,3).
Hormon uygulaması yapılan gruplara ait ovaryum dokusunda
PCNA immünreaktivitesinin geliĢen folliküllerde oosit çekirdeği düzeyinde
kuvvetli olduğu, buna karĢın sekonder follikülden itibaren oosit çekirdek
boyanmasının azaldığı belirlendi. Tüm folliküllerde granüloza ve teka
interna hücrelerinde yaygın tutulum dikkati çekiyordu. Ġmmün tepkime
çekirdek düzeyinde fazla sitoplazmik düzeyde ise daha azdı (Resim 4,5,6).
Hormon uygulaması yapılan gruba ait gebe ovaryumlarındaki
PCNA tutulumu tüm folliküllerde granüloza hücreleri, teka interna hücreleri
düzeyinde ve oosit çekirdeklerinde oldukça belirgindi. Ġmmünreaktivite
59
korpus luteum hücrelerinde ve atretik folliküllerdeki hücrelerde yaygındı.
Bazı stromal hücrelerde de oldukça kuvvetli PCNA boyanması dikkat
çekiciydi (Resim 7,8,9).
Klomifen
sitrat
uygulaması
yapılan
gruba
ait
kontrol
ovaryumlarında yapılan incelemelerde, PCNA tutulumunun hormon
uygulaması yapılan gruplara ait kontrol ovaryumlarına eĢdeĢ olduğu
görüldü (Resim 10,11).
Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokularında PCNA tutulumunun tüm folliküllerde, atretik olanlarda
stromada ve korpus luteumda son derece az düzeyde olduğu izlendi.
Granüloza hücrelerinde tepkime çekirdek düzeyinde ve az yoğundu.
Oositte ise belirgin bir tutulum izlenmiyordu. Buna karĢın çok sıralı primer
folliküllerde
birden
fazla
oositin
gözlemlenmesi
sitrat
uygulaması
dikkati
çekti
(Resim12,13,14).
Klomifen
yapılan
gruba
ait
gebe
ovaryumlarında PCNA immünreaktivitesinin, tüm folliküllerde granüloza ve
teka interna hücreleri düzeyinde olduğu ayırt ediliyordu. Oosit çekirdek ve
sitoplazmasında tutulum belirgindi. Ġmmünreaktivitenin korpus luteumda
daha az düzeyde olduğu belirlendi. Atretik folliküllerde de yaygın boyanma
dikkati çekti. GeliĢen folliküllerde bazen iki oositin varlığı ilgiyi çekiyordu
(Resim 15,16,17).
60
4.2.C-FOS BULGULARI:
Hormon
uygulaması
yapılan
gruba
ait
kontrol
grubu
ovaryumlarında c-FOS immünreaktivitesi değerlendirildiğinde, tutulumun
tüm folliküllerde oosit sitoplazması ile çekirdeğinde ve granüloza hücreleri
ile teka interna hücrelerinde olduğu görüldü. Boyanmanın çekirdek ve
sitoplazma düzeyinde granüler Ģekilde olduğu ve çoğunlukla orta dereceli
gözlemlendiği ilgiyi çekti. Bu gruba ait incelemelerde korpus luteum
hücrelerinde de orta dereceli immünreaktivite belirgindi (Resim 18,19,20).
Hormon uygulaması yapılan gruplara ait ovaryumlarda c-fos
immünreaktivitesi değerlendirildiğinde, tutulumun kontrole karĢın biraz
daha fazla ve çekirdek ile sitoplazma düzeyinde yaygın olduğu belirlendi
(Resim 21,22).
Hormon
uygulaması
yapılan
gruplara
ait
gebe
ovaryumlarında ise c-fos tutulumunun diğer gruplara karĢın biraz daha
özgün olduğu, sitoplazma ve çekirdekte granüller boyanmanın varlığı
dikkati çekti (Resim 23,24,25,26).
Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryumlarındaki c-fos immünreaktivitesi hormon grubu kontrolleri ile
eĢdeĢ yapı sergiliyordu (Resim 27,28,29).
Klomifen
sitrat
uygulaması
yapılan
grup
ile
hormon
uygulaması yapılan grup ovaryumları karĢılaĢtırıldığında, klomifen sitrat
61
uygulaması yapılan grupta c-fos tutulumunun ağırlıklı olarak teka interna
hücreleri ve stromada yoğunlaĢtığı izlendi. Ġmmün tepkimenin bu grupta,
tüm folliküllerde, granüloza hücreleri ve oositte, hormon grubuna karĢın az
olması ilgiyi çekti. Korpus luteum hücrelerinde de boyanma biraz daha
yoğundu (Resim 30,31,32).
Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokularında çok sıralı antrumun oluĢmaya baĢladığı folliküllerde c-fos
tutulumunun belirgin olarak arttığı ve granüloza hücrelerinde, oosit ile teka
interna hücrelerinde yaygınlaĢtığı belirlendi. Boyanma sitoplazmada
granüler Ģekildeydi. Ġmmünreaktivitenin atretik follikülde de belirginleĢtiği
ayırt edildi (Resim 33,34,35).
4.3.LIF BULGULARI:
Hormon uygulaması yapılan gruba ait kontrol ovaryumlarında
LIF tutulumu değerlendirildiğinde, korpus luteum, atretik ve geliĢen
folliküllerde ortadan zayıfa değiĢen tepkime gözlemlendi (Resim 36,37).
Hormon uygulaması yapılan gruba ait ovaryum dokusunda
yapılan incelemelerde LIF immünreaktivitesinin, kontrollere eĢdeĢ olduğu
dikkati çekiyordu (Resim 38,39,40).
Hormon uygulaması yapılan gruba ait gebe ovaryum
dokularında, bazı folliküllerde, granüloza, teka interna ve atretik follikül
hücrelerinde diğer gruplara karĢın tutulumun biraz daha yaygın olduğu
ilgiyi çekti (Resim 41,42,43).
62
Klomifen sitrat uygulaması yapılan gruba ait kontrol ovaryum
dokusu incelemelerinde LIF tutulumunun, hormon uygulaması yapılan
grubun kontrol grubuyla eĢdeĢ olduğu görüldü (Resim 44,45).
Klomifen sitrat uygulaması yapılan gruba ait ovaryumlardaki
değerlendirmelerde, LIF immünreaktivitesinin korpus luteum hücrelerinde
biraz daha fazla olduğu dikkati çekerken, diğer yapıların kontrol
grubundakilerle aynı görünümde olduğu belirlendi. Sekonder folliküllerde
ise oositi çevreleyen ilk sıra follikül hücrelerinde tutulumun biraz daha
arttığı görüldü (Resim 46,47).
Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokusu yapılan incelemelerinde, LIF immünreaktivitesi diğer gruplara
oranla artmıĢtı. Tutulum oosit sitoplazmasında az, granüloza ve teka
interna hücrelerinde ise biraz daha fazlaydı. Korpus luteumda da
boyanmanın orta dereceli ve yaygın olduğu dikkati çekti (Resim 48,49,50).
4.4.İSTATİSTİKSEL BULGULAR
Yapılan
istatistiksel
değerlendirmede,
klomifen
sitrat
uygulaması yapılan grupta klomifen sitrat uygulaması yapılan hayvanların
Graaf değerleri kontrol ve gebe grubuna karĢın farklıydı ve bu fark
anlamlıydı (p=0.006).
Hormon uygulaması yapılan deneklerin Graaf değerlerinde,
kontrol ve gebe gruplarına karĢın anlamlı fark belirlenemedi (p=0.095).
63
Hormon uygulaması yapılan kontrol grupları, klomifen sitrat
uygulaması yapılan kontrol grupları (p=0.749) ve hormon gebe ile klomifen
sitrat uygulaması yapılan gebe grupları (p=0.522) arasında istatistiksel
açıdan anlamlı bir fark yoktu. Ancak hormon uygulaması yapılan
deneklerle klomifen sitrat uygulaması yapılan deneklerin (p=0.006) Graaf
çapları anlamlı olarak birbirinden farklıydı.
Hormon ve klomifen sitrat uygulaması yapılan gruplar
değerlendirildiğinde, gruplar arası anlamlı bir fark bulunamadı (p=0.180,
p=0.565).
Hormon ve klomifen sitrat uygulaması yapılan gruplar
karĢılaĢtırıldığında ise anlamlı bir fark belirlenemedi (kontrol için p=1;
hormon-klomifen sitrat için p=0.121, gebeler için p=0.439).
Hormon ve klomifen sitrat uygulaması yapılan gruplar
arasında primordiyal follikül sayısında anlamlı bir fark yoktu (p= 0.354 ve
p= 0.802).
Hormon ve klomifen sitrat uygulaması yapılan gruplar
karĢılaĢtırıldığında ise anlamlı bir fark belirlenemedi (kontrol için p=0.936;
hormon-klomifen için p=0.936, gebeler için p=0.575).
Hormon ve klomifen sitrat uygulaması yapılan gruplar
arasında sekonder follikül sayısında anlamlı bir fark görülmedi (p= 0.795
ve p= 0.582).
64
Hormon ve klomifen sitrat uygulaması yapılan gruplar
karĢılaĢtırıldığında ise yine anlamlı bir fark belirlenemedi (kontrol için
p=0.422; hormon-klomifen sitrat için p=0.258, gebe için p=0.808).
65
RESİMLER
Resim 1: Hormon uygulaması yapılan gruplara ait kontrol ovaryum
dokusunda gelişim aşamalarındaki folliküller ve oositlerdeki PCNA
tutulumu izleniyor. (ÇKP): Çok sıralı primer follikül, (SF): Sekonder follikül,
(Ç): Çekirdek, (O): Oosit, (An): Antrum, (G): Granüloza hücreleri, (Tİ):
Teka interna hücreleri (İmmünperoksidaz-HematoksilenX100).
66
Resim 2A,B: Hormon uygulaması yapılan gruplara ait kontrol ovaryum
dokusunda; gelişen foliküllerde oosit çekirdeği düzeyinde kuvvetli olmakla
birlikte, sekonder folikülden itibaren azalan PCNA immünreaktivitesi
izleniyor. (ÇKP): Çok sıralı primer follikül, (TKP): Tek sıralı primer follikül,
(Ç): Çekirdek, (O): Oosit, (G): Granüloza hücreleri (İmmünperoksidazHematoksilen AX400, BX400).
67
Resim 3: Hormon uygulaması yapılan gruplara ait kontrol ovaryum
dokusunda, antrumu belirgin sekonder folliküllerde oositteki PCNA
tutulumunun belirgin olarak azaldığı görülüyor. (O): Oosit, (An): Antrum,
(G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (ZP): Zona pellusida
(İmmünperoksidaz-Hematoksilen X400).
68
Resim 4A,B,C: Hormon uygulaması yapılan gruplara ait ovaryum
dokusunda sekonder folliküllerde oosit çekirdek tutulumunun diğer
folliküllere karşın az olduğu görülüyor. (SF): Sekonder follikül, (O): Oosit,
(An): Antrum, (G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (ZP):
Zona pellusida (İmmünperoksidaz-Hematoksilen AX100, BX100, CX100).
69
Resim
5A,B:
Hormon
uygulaması yapılan
gruplara
ait ovaryum
dokusunda PCNA immünreaktivitesinin özellikle çekirdek düzeyinde ve
granüloza hücrelerinde yaygın olduğu dikkati çekiyor. (TKP): Tek sıralı
primer follikül, (Ç): Çekirdek, (O): Oosit, (G): Granüloza hücreleri
(İmmünperoksidaz-Hematoksilen AX400, BX400).
70
Resim
6A,B:
Hormon
uygulaması
yapılan
gruplara
ait ovaryum
dokusunda yaygın PCNA immünreaktivitesi görülüyor. (O): Oosit, (An):
Antrum, (G): Granüloza hücreleri, (ZP): Zona pellusida (İmmünperoksidazHematoksilen AX400, BX400).
71
Resim
7A,B,C,D:
Hormon
uygulaması
yapılan
gruba
ait
gebe
ovaryumlarında çeşitli gelişim evrelerindeki folliküller ve korpus luteumda
PCNA immünreaktivitesi görülüyor. (SF): Sekonder follikül, (O): Oosit,
(An): Antrum, (G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (ZP):
Zona pellusida, (ÇKP): Çok sıralı primer follikül, (Ç): Çekirdek, (KL):
Korpus luteum (İmmünperoksidaz-Hematoksilen AX100, BX100, CX100,
DX100).
72
Resim 8A,B: Hormon uygulaması yapılan gruba ait gebe ovaryumlarında
PCNA immünreaktivitesinin çekirdek düzeyinde son derece belirgin olduğu
görülüyor. (ÇKP): Çok sıralı primer follikül, (Ç): Çekirdek, (O): Oosit, (G):
Granüloza hücreleri, (Tİ): Teka interna hücreleri (İmmünperoksidazHematoksilen AX400, BX400).
73
Resim 9: Hormon uygulaması yapılan gruba ait gebe ovaryumlarında
PCNA boyamasında; oosit çekirdeğinde ve granuloza hücrelerinde belirgin
tutulum izleniyor. (SF): Sekonder follikül, (An): Antrum, (G): Granüloza
hücreleri,
(Tİ):
Teka
interna
hücreleri,
(ZP):
Zona
pellusida
(İmmünperoksidaz-Hematoksilen X400).
74
Resim 10A,B: Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryum dokusunda PCNA tutulumunun doku genelinde hormon kontrol
grubuna eşdeş olduğu görülüyor. (SF): Sekonder follikül, (An): Antrum,
(G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (ZP): Zona pellusida,
(O): Oosit (İmmünperoksidaz-Hematoksilen AX100,BX100).
75
Resim 11A,B: Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryum dokusunda yapılan PCNA boyamasında; çekirdek tutulumunun
sekonder folikülde primer foliküllere karşın kuvvetli olduğu ayırt ediliyor.
(SF): Sekonder follikül, (An): Antrum, (G): Granüloza hücreleri, (Tİ): Teka
interna hücreleri, (ZP): Zona pellusida, (O): Oosit, (Ç): Çekirdek, (TKP):
Tek sıralı primer follikül (İmmünperoksidaz-Hematoksilen AX400,BX400).
76
Resim 12: Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokularında PCNA immünreaktivitesinin tüm folliküllerde, atretik olanlarda
stromada ve korpus luteumda son derece az düzeyde olduğu dikkati
çekiyor. (ÇKP): Çok sıralı primer follikül, (SF): Sekonder follikül, (AF):
Atretik follikül, (S): Stroma, (KL): Korpus luteum,
(Ç): Çekirdek
(İmmünperoksidaz-Hematoksilen X100).
77
Resim 13A,B: Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokularında büyük büyültmeli resimlerde, granüloza hücrelerinde ve
çekirdek düzeyinde PCNA tutulumunun az olduğu görülürken, oositte
belirgin bir tutulum izlenmiyor. (ÇKP): Çok sıralı primer follikül, (Ç):
Çekirdek, (PF): Primordiyal follikül, (G): Granüloza hücreleri, (O): Oosit
(İmmünperoksidaz-Hematoksilen AX400, BX400).
78
Resim 14A,B: Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokularında PCNA boyamasında; çok sıralı primer folliküllerde birden fazla
oositin varlığı dikkati çekiyor. (SF): Sekonder follikül, (An): Antrum(An),
(G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (O): Oosit, (Ç):
Çekirdek, (ÇKP): Çok sıralı primer follikül (İmmünperoksidaz-Hematoksilen
AX400, BX400).
79
Resim 15A,B,C: Klomifen sitrat uygulaması yapılan gruba ait gebe
ovaryumlarında PCNA boyamasında tutulumun tüm folliküllerde granüloza
ve teka interna hücreleri düzeyinde olduğu ayırt ediliyordu. (An): Antrum,
(G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (O): Oosit, (Ç):
Çekirdek, (ÇKP): Çok sıralı primer follikül, (KL): Korpus luteum, (PF):
Primordiyal
follikül
(İmmünperoksidaz-Hematoksilen
AX100,
BX100,
CX100).
80
Resim 16A,B,C: Klomifen sitrat uygulaması yapılan gruba ait gebe
ovaryumlarında PCNA immünreaktivitesinin özellikle çekirdek düzeyinde
belirgin olduğu görülürken, gelişen foliküllerde iki oositin varlığı ayırt
ediliyor. (O): Oosit, (Ç): Çekirdek, (TKP): Tek sıralı primer follikül
(İmmünperoksidaz-Hematoksilen AX400, BX400, CX400).
81
Resim 17A,B: Klomifen sitrat uygulaması yapılan gruba ait gebe
ovaryumlarında PCNA boyamasında tutulumun oosit, granuloza ve teka
interna hücrelerinde belirgin olduğu görülüyor. (SF): Sekonder follikül,
(An): Antrum, (G): Granüloza hücreleri, teka interna hücreleri(Tİ), oosit(O),
çekirdek(Ç),
çok
sıralı
primer
follikül(ÇKP),
zona
pellusida(ZP).
(İmmünperoksidaz-Hematoksilen AX400, BX400).
82
Resim 18: Hormon uygulaması yapılan gruba ait kontrol grubu
ovaryumlarında c-FOS tutulumunun, tüm folliküllerde oosit sitoplazması,
granüloza hücreleri ve teka interna hücrelerinde olduğu görülüyor. (ÇKP):
Çok sıralı primer follikül, (SF): Sekonder follikül, (An) Antrum, (G):
Granüloza hücreleri, (O): Oosit (İmmünperoksidaz-Hematoksilen X100).
83
Resim 19A,B: Hormon uygulaması yapılan gruba ait kontrol grubu
ovaryumlarında c-FOS immünreaktivitesinin çekirdek ve sitoplazma
düzeyinde, granüler şekilde ve çoğunlukla orta dereceli olduğu ilgiyi
çekiyor. (SF): Sekonder follikül, (An): Antrum, (G): Granüloza hücreleri,
(Tİ): Teka interna hücreleri, (O): Oosit, (Ç): Çekirdek, (ÇKP): Çok sıralı
primer follikül, (ZP): Zona pellusida (İmmünperoksidaz-Hematoksilen
AX400, BX400).
84
Resim 20: Hormon uygulaması yapılan gruba ait kontrol grubu
ovaryumlarında c-FOS boyamasında daha büyük büyültmelerde de
tutulumun granüler şekilde ve orta dereceli olduğu izleniyor. (G):
Granüloza hücreleri, (O): Oosit, (Ç): Çekirdek (İmmünperoksidazHematoksilen X400).
85
Resim 21A,B: Hormon uygulaması yapılan gruba ait ovaryum dokusunda
c-FOS boyamasında tutulumun bir önceki gruba karşın doku genelinde
biraz daha arttığı görülüyor. (SF): Sekonder follikül, (An): Antrum, (G):
Granüloza hücreleri, (Tİ): Teka interna hücreleri, (O): Oosit, (Ç): Çekirdek,
(ÇKP): Çok sıralı primer follikül, (ZP): Zona pellusida (İmmünperoksidazHematoksilen AX100, BX100).
86
Resim 22A,B: Hormon uygulaması yapılan gruba ait ovaryum dokusunda
c-FOS immünreaktvitesinin daha büyük büyültmelerde çekirdek ile
sitoplazma düzeyinde yaygın olduğu ayırt ediliyor. (SF): Sekonder follikül,
(An): Antrum, (G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (O):
Oosit, (Ç): Çekirdek, (ZP): Zona pellusida, (ÇKP): Çok sıralı primer follikül
(İmmünperoksidaz-Hematoksilen AX400, BX400).
87
Resim 23A,B: Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda c-FOS immünreaktivitesinin diğer gruplara karşın daha özgün
olduğu dikkati çekiyor. (SF): Sekonder follikül, (An): Antrum, (G):
Granüloza hücreleri, (Tİ): Teka interna hücreleri, (O): Oosit, (Ç): Çekirdek,
(ÇKP): Çok sıralı primer follikül, (ZP): Zona pellusida, (AF): Atretik follikül
(İmmünperoksidaz-Hematoksilen AX100, BX100).
88
Resim 24: :
Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda c-FOS tutulumunun çekirdek ve sitoplazmada granüler halde
olduğu izleniyor. (ÇKP): Çok sıralı primer follikül, (G): Granüloza hücreleri,
(O): Oosit, (PF): Primordiyal follikül (İmmünperoksidaz-Hematoksilen
X400).
89
Resim 25: Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda c-FOS boyamasında, sekonder folllikülde oosit ve granuloza
hücrelerinde belirgin tutulum izleniyor. (SF): Sekonder follikül, (An):
Antrum, (G): Granüloza hücreleri, (Tİ): Teka interna hücreleri, (O): Oosit,
(ÇKP): Çok sıralı primer follikül (İmmünperoksidaz-Hematoksilen X400).
90
Resim 26: Hormon uygulaması yapılan gruplara ait gebe ovaryum
dokusunda daha büyük büyültmelerde, çekirdek ve oosit düzeyinde
belirgin c-fos tutulumu görülüyor. (SF): Sekonder follikül, (An): Antrum,
(G): Granüloza hücreleri, (O): Oosit, (Ç): Çekirdek, (ZP): Zona pellusida
(İmmünperoksidaz-Hematoksilen X400).
91
Resim 27: Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryum dokusundaki c-FOS imünreaktivitesinin doku genelinde hormon
kontrol grubuna eşdeş olduğu izleniyor. (SF): Sekonder follikül, (Ç):
Çekirdek, (O): Oosit (İmmünperoksidaz-Hematoksilen X100).
92
Resim 28: Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryum dokusunda yapılan c-FOS boyamasında tutulumun çekirdek ve
sitoplazma düzeyinde çoğunlukla orta dereceli gözlemlenmesi ilgiyi
çekiyor. (O): Oosit, (ÇKP): Çok sıralı primer follikül, (G): Granüloza
hücreleri (İmmünperoksidaz-Hematoksilen X400).
93
Resim 29: Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryum dokusundaki c-FOS immünreaktivitesi daha büyük büyültmelerde
incelendiğinde tutulumun oositte yaygın ve granüler şekilde olduğu
görülüyor. (SF): Sekonder follikül, (An(: Antrum, (G): Granüloza hücreleri,
(O): Oosit, (ZP): Zona pellusida (İmmünperoksidaz-Hematoksilen X400)
94
Resim 30A,B: Klomifen sitrat uygulaması yapılan gruba ait ovaryum
dokusunda c-FOS immünreaktivitesinin doku genelinde stromada daha
belirgin olduğu görülüyor. (SF): Sekonder follikül, (An): Antrum, (G):
Granüloza hücreleri, (O): Oosit, (Ç): Çekirdek, (ZP): Zona pellusida,
(ÇKP):
Çok
sıralı
primer
follikül,
(Tİ):
Teka
interna
hücreleri
(İmmünperoksidaz-Hematoksilen AX100, BX100).
95
Resim 31: Klomifen sitrat uygulaması yapılan grup ovaryum dokusunda
yapılan c-FOS boyamasında granuloza hücrelerinde tutulumun az yoğun
olduğu görülüyor. (SF): Sekonder follikül, (An): Antrum, (G): Granüloza
hücreleri, (O): Oosit, (Ç): Çekirdek, (ZP): Zona pellusida, (ÇKP): Çok sıralı
primer follikül, (Tİ): Teka interna hücreleri, (TKP): Tek sıralı primer follikül,
(PF): Primordiyal follikül (İmmünperoksidaz-Hematoksilen X400).
96
Resim 32: : Klomifen sitrat uygulaması yapılan grup ovaryum dokusunda
yapılan c-FOS boyamasında tutulumun hormon uygulaması yapılan gruba
karşın az olduğu izleniyor. (O): Oosit, (G): Granüloza hücreleri, (An):
Antrum (İmmünperoksidaz-Hematoksilen X400).
97
Resim 33A,B: Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokusunda, doku genelinde folliküllerde c-fos tutulumunun belirgin olarak
arttığı ve granüloza hücrelerinde, oosit ile teka interna hücrelerinde
yaygınlaştığı ayırt ediliyor. (SF): Sekonder follikül, (An): Antrum, (G):
Granüloza hücreleri, (O): Oosit, (Ç): Çekirdek, (ZP): Zona pellusida, (Tİ):
Teka interna hücreleri, (ÇKP): Çok sıralı primer follikül (İmmünperoksidazHematoksilen AX100, BX100).
98
Resim 34: Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokusunda yapılan c-FOS boyamasında tutulumun stromada belirginleştiği
görülüyor. (ÇKP): Çok sıralı primer follikül, (G): Granüloza hücreleri, (O):
Oosit, (Ç): Çekirdek (İmmünperoksidaz-Hematoksilen X400).
99
Resim 35A,B: Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokusunda yapılan c-FOS boyamasında tutulumun sitoplazmada ve
granüler şekilde olduğu görülüyor. (SF): Sekonder follikül, (An): Antrum,
(G): Granüloza hücreleri, (O): Oosit, (Ç): Çekirdek, (ZP): Zona pellusida
(İmmünperoksidaz-Hematoksilen AX400, BX400).
100
Resim 36: Hormon uygulaması yapılan gruba ait kontrol ovaryum
dokusunda yapılan LIF boyamasında korpus luteum, atretik ve gelişen
folliküllerde ortadan zayıfa değişen tutulum gözlemleniyor. (SF): Sekonder
follikül, (An): Antrum, (G): Granüloza hücreleri, (O): Oosit, (AF): Atretik
follikül, (KL): Korpus luteum (İmmünperoksidaz-Hematoksilen X100).
101
Resim 37A,B: Hormon uygulaması yapılan gruba ait kontrol ovaryum
dokusunda yapılan LIF boyamasında büyük büyültmelerde foliküllerde
zayıftan ortaya değişen tutulum izleniyor. (SF): Sekonder follikül, (An):
Antrum, (G): Granüloza hücreleri, (O): Oosit, (ÇKP): Çok sıralı primer
follikül (İmmünperoksidaz-Hematoksilen AX400, BX400).
102
Resim 38: Hormon uygulaması yapılan gruba ait ovaryum dokusunda
yapılan LIF boyamasında; immünreaktivitenin kontrol grubuna eşdeş
olduğu dikkati çekiyor. (SF): Sekonder follikül, (ÇKP): Çok sıralı primer
follikül, (KL): Korpus luteum (İmmünperoksidaz-Hematoksilen X40).
103
Resim 39: : Hormon uygulaması yapılan gruba ait ovaryum dokusunda
yapılan LIF boyamasında; tek ve çok sıralı primer foliküllerde tutulumun
zayıf-orta dereceli olduğu görülüyor. (TKP): Tek sıralı primer follikül,
(ÇKP): Çok sıralı primer follikül, (O): Oosit, (G): Granüloza hücreleri
(İmmünperoksidaz-Hematoksilen X400).
104
Resim 40: Hormon uygulaması yapılan gruba ait ovaryum dokusunda
büyük büyültmelerde oosit ve çekirdekteki zayıf LIF tutulumu izleniyor
(An): Antrum, (G): Granüloza hücreleri, (O): Oosit, (Ç): Çekirdek
(İmmünperoksidaz-Hematoksilen X400).
105
Resim 41: Hormon uygulaması yapılan gruba ait gebe grubu ovaryum
dokusunda yapılan LIF boyamasında, doku genelinde bazı folliküllerde
diğer gruplara karşın tutulumun biraz daha yaygın olduğu ilgiyi çekiyor.
(SF): Sekonder follikül, (ÇKP): Çok sıralı primer follikül, (KL): Korpus
luteum (İmmünperoksidaz-Hematoksilen X40).
106
Resim 42A,B: Hormon uygulaması yapılan gruba ait gebe ovaryum
dokusunda yapılan LIF boyamasında; tek ve çok sıralı primer foliküllerde
bir önceki gruba karşın tutulumun biraz daha arttığı görülüyor. (TKP): Tek
sıralı primer follikül, (SF): Sekonder follikül, (O): Oosit, (Ç): Çekirdek, (G):
Granüloza hücreleri, (Tİ): Teka interna hücreleri (İmmünperoksidazHematoksilen AX400, BX400).
107
Resim 43: Hormon uygulaması yapılan gruba ait gebe ovaryum
dokusunda sekonder folikülde ve oositteki LIF immünreaktivitesi izleniyor.
(SF): Sekonder follikül, (An): Antrum, (G): Granüloza hücreleri, (O): Oosit,
(Tİ): Teka interna hücreleri (İmmünperoksidaz-Hematoksilen X400).
108
Resim 44A,B: Klomifen sitrat uygulaması yapılan gruba ait kontrol
ovaryum dokusunda yapılan LIF boyamasında
tutulumun hormon
uygulaması yapılan grubun kontrol grubuyla eşdeş olduğu görülüyor. (SF):
Sekonder follikül, (An): Antrum, (G): Granüloza hücreleri, (O): Oosit,
(ÇKP): Çok sıralı primer follikül, (KL): Korpus luteum (İmmünperoksidazHematoksilen AX100, BX100).
109
Resim 45A,B,C:
Klomifen sitrat uygulaması yapılan gruba ait kontrol
ovaryum dokusunda; primordiyal, çok sıralı primer ve sekonder foliküllerde
LIF immünreaktivitesinin son derece zayıf olduğu izleniyor. (SF): Sekonder
follikül, (An): Antrum, (G): Granüloza hücreleri, (O): Oosit, (ÇKP): Çok
sıralı
primer
follikül,
(PF):
Primordiyal
follikül
(İmmünperoksidaz-
Hematoksilen AX400, BX400, CX400).
110
Resim 46A,B: Klomifen sitrat uygulaması yapılan gruplara ait ovaryum
dokusunda, LIF immünreaktivitesinin korpus luteum hücrelerinde biraz
daha çok olduğu dikkati çekiyor. (SF): Sekonder follikül, (An): Antrum, (G):
Granüloza hücreleri, (O): Oosit, (ÇKP): Çok sıralı primer follikül, (TKP):
Tek sıralı primer follikül, (KL): Korpus luteum, (AF): Atretik follikül
(İmmünperoksidaz-Hematoksilen AX100, BX100).
111
Resim 47A,B: Klomifen sitrat uygulaması yapılan gruplara ait ovaryum
dokusunda yapılan LIF boyamasında, sekonder folliküllerde oositi
çevreleyen ilk sıra hücrelerde tutulumun biraz daha arttığı izleniyor. (SF):
Sekonder follikül, (An): Antrum, (G): Granüloza hücreleri, (O): Oosit, (Ç):
Çekirdek, (ÇKP): Çok sıralı primer follikül, (TKP): Tek sıralı primer follikül
(İmmünperoksidaz-Hematoksilen AX400, BX400).
112
Resim 48A,B: Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokusunda LIF immünreaktivitesinin diğer gruplara karşın artmış olması
ilgiyi çekiyor. (SF): Sekonder follikül, (An): Antrum, (G): Granüloza
hücreleri, (O): Oosit, (KL): Korpus luteum (İmmünperoksidaz-Hematoksilen
AX100, BX100)
113
Resim 49: : Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokusunda LIF tutulumunun oosit sitoplazmasında az yoğun olduğu
izleniyor. (ÇKP): Çok sıralı primer follikül, (G): Granüloza hücreleri, (O):
Oosit, (Tİ): Teka interna hücreleri (İmmünperoksidaz-Hematoksilen X400).
114
Resim 50: Klomifen sitrat uygulaması yapılan gruba ait gebe ovaryum
dokusunda; sekonder folikülde granüloza hücrelerinde ve teka interna
hücrelerinde LIF immünreaktivitesinin, oosite karşın biraz daha fazla
olduğu görülüyor. (An): Antrum, (G): Granüloza hücreleri, (O): Oosit, (Tİ):
Teka interna hücreleri (İmmünperoksidaz-Hematoksilen X400).
115
Tablo 1. Hormon ve klomifen grubuna ait Graaf değerlerinin istatistiksel açıdan
değerlendirilmesi
Uygulama
Hormon
Hormon
Hormon
(Gebe)
(Kontrol)
p=0.095
p=0.095
Klomifen
Klomifen
(Gebe)
p=0.006
Hormon (Gebe)
p=0.522
Klomifen (Kontrol)
p=0.749
p=0.006
p=0.006
Tablo 2. Hormon ve klomifen sitrat uygulaması yapılan gruplara ait ovaryum çaplarının
istatistiksel açıdan değerlendirilmesi
Uygulama
Hormon
Hormon
Hormon
(Gebe)
(Kontrol)
p=0.180
p=0.180
Klomifen
(Gebe)
p=0.121
Hormon (Gebe)
Klomifen (Kontrol)
Klomifen
p=0.439
p=1.0
p=0.565
p=0.565
116
Tablo 3. Hormon ve klomifen sitrat uygulaması yapılan gruplara ait primordiyal follikül
sayılarının istatistiksel açıdan değerlendirilmesi
Uygulama
Hormon
Hormon
Hormon
(Gebe)
(Kontrol)
p=0.354
p=0.354
Klomifen
Klomifen
(Gebe)
p=0.936
Hormon (Gebe)
p=0.575
Klomifen (Kontrol)
p=0.936
p=0.802
p=0.802
Tablo 4. Hormon ve klomifen sitrat uygulaması yapılan gruplara ait sekonder follikül
sayılarının istatistiksel açıdan değerlendirilmesi
Uygulama
Hormon
Hormon Hormon
(Gebe)
(Kontrol)
p=0.795
p=0.795
Klomifen Klomifen
(Gebe)
p=0.258
Hormon (Gebe)
Klomifen
p=0.808
p=0.422
p=0.582
p=0.582
(Kontrol)
117
Tablo 5. Hormon uygulaması yapılan gruba ait tanıtıcı istatistikler
Hormon
Hormon gebe
Hormon kontrol
P. fol
S fol.
P. fol
S fol.
P. fol
S fol.
Ortalama
2.82
5.35
4.81
3.46
4.75
3.51
St. Sapma
1.63
4.05
2.93
2.28
2.79
2.12
Ortanca
2.5
4.0
4.0
3.0
4.0
1
Minimum
1
1
1
1
1
10
Maksimum
7
17
13
10
12
3
Tablo 6. Klomifen sitrat uygulaması yapılan grubuna ait tanıtıcı istatistikler
Klomifen
Klomifen gebe
klomifen kontrol
P. fol
S fol.
P. fol
S fol.
P. fol
S fol.
Ortalama
4.77
3.51
5.27
3.42
4.72
3.69
St. Sapma
2.97
1.20
3.88
1.91
2.82
2.14
Ortanca
4.0
3.0
4.0
3.0
4.0
3.0
Minimum
1
1
2
1
1
1
Maksimum
12
7
4
8
13
10
118
Tablo 8. Follikül sayılarına ait tanıtıcı istatistikler
Hormon
Hormon
Hormon
gebe
kont
Klomifen
Klomifen
Klomifen
gebe
kontrol
Ortalama
4469.6
4013.3
3861.0
3459.7
3403.0
3831.0
St. Sapma
705.1
770.1
550.9
954.5
1081.3
550.9
Ortanca
4503.8
4148.6
3909.9
3396.5
3588.0
3909.9
Minimum
2295.6
1673.2
2689.4
1487.8
1248.8
2913.8
Maksimum
5650.8
5466.5
4843.6
5176.4
6585.3
4646.1
119
5.TARTIŞMA
Herhangi bir doğum kontrol yöntemi uygulamadan, haftada
en az iki kez düzenli bir cinsel yaĢama karĢın, bir yıl süreyle gebelik
oluĢmamasına infertilite denir. Ġnfertilite sorunu üreme çağındaki çiftlerin
yaklaĢık %10-15‘ini etkilemektedir. Doğum kontrol yöntemi kullanmayan
normal çiftlerin %57‘si ilk 3 ayda, %72‘si ilk 6 ay içinde, %85‘i bir yıl içinde
ve %93‘ü ilk 2 yıl içinde gebe kalabilmektedir. Erkek faktörü infertilitenin,
%35‘ini oluĢturur. Kadın infertilitesinde ise en sık nedenler; %15 oranında
ovulasyon bozukluğu, %40 tubal patoloji, %10‘ dan az olarak anatomik
bozukluklardır. Geri kalanı ise nedeni açıklanamamıĢ infertilite olgularıdır.
1960‘lı yılların baĢlarına dek anovulasyon ve diğer infertilite tiplerinin etkili
tedavisi tam olarak bilinmiyordu. 1961 yılında Greenblatt ve arkadaĢları ilk
olarak o zamanki adıyla MRL-41 olarak bilinen klomifen sitrat kullanarak
infertil olgularda baĢarıya ulaĢmıĢlardır. 31,32,35
Gemzell ve arkadaĢları 1958 yılında insan hipofiz bezi
gonadotropinlerini kullanarak ovulasyon indüksiyonu yapmaya baĢlamıĢlar
ve 1960 yılında gebelik elde etmiĢlerdir. Gemzell ve Bettendorf
hipofizektomili hastalara insan hipofiz bezi gonadotropini uygulayarak
gebelik
sağlamıĢlardır.
1970‘li
yıllarda
anovulasyon
tedavisi
için
gonadotropin salgılatıcı hormon (GnRH) ve prolaktin baskılayıcı ajanlar
olarak iki seçenek öne sürülmüĢtür. GnRH izole edilmiĢ ve yapısı
tanımlanmıĢtır. Kısa bir süre sonra laboratuvarda doğal GnRH preparat
sentezi ile klinik kullanım için daha uygun hale gelmiĢtir. GnRH tedavisinin,
hipotalamik kökenli hipogonadotropik amenoreli kadınlarda ovulasyon
indüksiyonu ve gebelik gerçekleĢmesinde çok etkili olduğu kanıtlanmıĢtır.
36,37,38,39
120
Günümüzde ovulasyon indüksiyonunda amaç, anovulatuar
hastalarda ovulasyonu baĢlatmaktır. Ovulasyon sağlanabildiği koĢullarda
ise esas amaç tek follikül ovulasyonudur. Ovulasyon indüksiyonu için
kullanılan gonadotropinler 3 türdür. Bunlar; üriner, saf üriner ve
rekombinant gonadotropinlerdir. Ġnsan menapozal gonadotropini (hMG,
menotropin) yaklaĢık 30 yıl boyunca kullanılmıĢtır. Yeni hMG preparatları
ise eskilerinden daha iyi arılaĢtırılmıĢ ve cilt altı uygulanabilen formlardır.
15,40
Gonadotropinlerin ovulasyon indüksiyonunda yüksek doz ve
uzun süreli kullanımının ovaryum kanserini tetiklediği bilinmektedir.
Gonadotropinler siklojenaz 2 ‗yi yükseltip prostoglandin üretimini artırırlar.
Bu epitel bazal membranlarında kayıplara neden olur ve epitel hücre
biyolojisini, hücre hücre sinyal iletimini ve düzenleniĢini değiĢtirir. 115
Yüksek düzeydeki gonadotropinler östrojen üretimine neden
olurlar. Östrojen öncülleri ovaryum yüzey epitelinde inklüzyon kistleri
oluĢmasını tetikler ve bu hasarlı epitel hücreleri çoğalır. Bunlar kansere
dönüĢümde esas rolü oynarlar. Nieto ve arkadaĢlarının yaptığı çalıĢmada
ovulasyon indüksiyonunda yüksek displazi oranları gözlenmiĢtir.
Yapılan
çalıĢmalarda
ovaryum
displazisinin
116,117
sınırdaki
ovaryum kanserleri ile ortak yapısal bulgular içerdiği belirlenmiĢtir. Birçok
çalıĢmada da ovulasyon indüksiyonunun sınırdaki ovaryum kanser riskini
artırdığı bildirilmiĢtir. 118,119,120
121
Chene ve arkadaĢları ovulasyon indüksiyonu uygulanmıĢ
hastalarla yaptıkları çalıĢmalarda, ovulasyon indüksiyonu ilaçlarının,
yüksek doz ve uzun süreli kullanımda ovaryum epitel displazisi
oluĢturduğunu belirtmiĢlerdir. 121
Çelik ve arkadaĢları çalıĢmalarında ovaryum displazisini 3
grupta
incelemiĢlerdir.
Sırasıyla
1,
3,
6
gonadotropin
döngüsü
uygulamıĢlar ve ovulasyon döngüsü arttıkça displazinin de arttığını
saptamıĢlardır. 122
Nieto ve arkadaĢlarının yaptığı bir diğer çalıĢmada, hormonal
etiyolojinin displazik olguları artırdığı bildirilmiĢtir. Üreme ilaçları ve
sınırdaki ovaryum kanseri arasında olası bir iliĢki olabileceğine dikkat
çekilmiĢtir. Literatürdeki diğer çalıĢmalarda da üreme ilaçlarından klomifen
sitratın borderline ovaryum tümörlerini artırıcı etkisi olduğu ve tedavi
sırasında bunun da göz önünde bulundurulması gerektiği vurgulanmıĢtır.
117,123,124,125,126
Tan ve arkadaĢları farelerle yaptıkları çalıĢmada, artmıĢ
ovulasyon sayısının ovaryum kist sayısını da artırabileceği varsayımı
üzerinde durmuĢlardır. ÇalıĢma için yaĢam süresine bağlı artan ovulasyon
sayıları için 1, 3, 8 ve 12 aylık fare grupları kullanılmıĢtır. Sonuçta 8 ve 12
aylık farelerde 1 ve 3 aylık farelere karĢın daha fazla ovaryum kistine
rastlanmıĢtır. AraĢtırıcılar kistlerin bening inklüzyon kistleri olduklarını
bildirmiĢlerdir. ÇalıĢmanın önemli sonuçlarından biri ovulasyonun ovaryum
yüzey epitelinden köken alan inklüzyon kistlerini artırabileceğidir. 127
122
Ovaryum aromatazlarının biyosentezi ve östrojen üretiminin
gonadotropinlerin ( FSH ve LH) etkisi altında olduğu bilinmektedir. Zhuang
ve arkadaĢları sıçan granuloza hücre kültürü ile yaptıkları çalıĢmada
östrojenlerin
(diethylstilbestrol
ve
17g-estradiol)
aromataz
erkini
çoğalttığını belirtmiĢlerdir. AraĢtırıcılar aynı çalıĢmada klomifen sitratın da
östrojen ile aynı etkiyi gösterdiğini saptamıĢlardır. Bu bulgulara dayanarak
ovaryum
folliküllerinin
östrojeni,
pozitif
geri-bildirim
düzeneği
ile
denetlediğini bildirmiĢlerdir. 128
Williams ve arkadaĢları 55 ÜYTE hastası ile yaptıkları
çalıĢmada kısa ve uzun tedavi protokollerini karĢılaĢtırmıĢlardır. Kısa
protokol uygulanacak gruptaki hastalara menstruel döngülerinin 3-7
günleri arasında klomifen sitrat, 9. günde ise gonadotropin (üriner FSH,
yüksek saflaĢtırılmıĢ FSH ve rekombinant FSH) uygulanmıĢtır. Follikül
çapı 16 mm‘yi geçene dek gonadotropin uygulanmaya devam edilmiĢ ve
folliküller bu büyüklüğe ulaĢtıklarında hCG uygulanmıĢtır. Uzun protokol
uygulanan hasta grubunda ise bir önceki döngünün 21. gününde GNRH
antagonisti uygulanmaya baĢlanmıĢ ve doz azaltılarak bir sonraki
döngünün 1. gününde kesilmiĢtir. 3. günde ise FSH uygulanmıĢtır. Sonraki
iĢlemler ise kısa protokoldeki gibi yapılmıĢtır. Sonuç olarak uzun protokol
uygulanan grupta E2 düzeyleri, oosit sayısı, implantasyon oranları, gebelik
oranlarının diğer gruba karĢın daha yüksek olduğu görülmüĢtür. Kısa
protokolün ise maliyet olarak daha uygun olduğunu ancak kaliteli embriyo
sayısının çokluğu ve embriyo dondurma iĢlemi için uzun protokolünün
yeğlenmesi gerektiğini bildirmiĢlerdir. 129
Hung Yu Ng ve arkadaĢları baĢarılı bir ÜYTE için; tedavi
öncesi yaĢ, bazal FSH yoğunluğu, ovaryum oylumu ve antral follikül
sayısını karĢılaĢtırarak ovaryumun tedaviye yanıtında hangisinin daha iyi
123
bir belirteç olduğunu araĢtırmıĢlardır. 128 kadın ile yapılan çalıĢmada
antral follikül sayısının en iyi belirteç olduğuna karar vermiĢlerdir. Antral
follikül sayısı çok olan hastalarda oositlerin sayısının da fazla olduğunu
belirlemiĢlerdir. Aynı çalıĢmada diğer belirtecin bazal FSH değerleri
olduğunu söylemiĢledir. Yüksek bazal FSH değerine sahip hastaların
tedavi sonrası az sayıda oosite sahip oldukları görülmüĢtür. Küçük
ovaryumların daha az antral follikül içermeleri nedeniyle büyük boyutlu
ovaryuma sahip hastalarda daha iyi yanıt alındığını bildirmiĢlerdir. Bir diğer
belirteç olan yaĢ ise en son sırada yer almıĢtır. YaĢ ile birlikte primordiyal
follikül sayısı azaldığı için tedavi sonrası ovaryumdaki oosit sayısı da
azalmıĢtır. 130
Lass
ve
arkadaĢları
çalıĢmalarında
küçük
boyutlu
ovaryumların HMG ile ovulasyon indüksiyonuna zayıf yanıt verdikleri
varsayımını araĢtırmıĢlardır. ÜYTE tedavisi gören 140 infertil hasta ile
yaptıkları çalıĢmada sonuç olarak ovaryum oylumu ve ovaryum rezervi
arasında güçlü bir iliĢki olduğunu saptamıĢlardır. Küçük ovaryumların
HMG‘ye zayıf yanıt verdiği ve tedavi iptali oranının yüksek olduğunu
bildirmiĢlerdir. Ġki grup arasında döllenme ve gebelik oranlarında bir fark
görülmemiĢtir. Ancak ovaryum boyutunun infertilite tedavisinde göz önüne
alınması gereken bir belirteç olduğuna dikkat çekmiĢlerdir.
131
Filicori ve arkadaĢları 1999 yılında yaptıkları çalıĢmada;
LH‘ın rolünü belirlemek için GnRH baskılanmıĢ hastaların bir grubuna 150
IU saflaĢtırılmıĢ FSH diğer grubuna ise 150 IU saf FSH‘ a ek olarak 50 IU
hCG vererek ovulasyon indüksiyonu uygulanmıĢtır. Sonuç olarak LH
erkinin FSH‘nin etkisini arttırdığı, follikül geliĢimini hızlandırdığı, ovulasyon
indüksiyonu süresini azalttığını ve FSH gereksinimini azaltarak maliyeti
düĢürdüğünü saptamıĢlardır.
132
124
Willis ve arkadaĢları tarafından yapılan çalıĢmada; farklı
boyutlardaki folliküllerin granüloza hücrelerinin FSH ve LH‘ ye yanıt olarak
östrojen ve progesteron üretimleri değerlendirilmiĢtir. ÇalıĢmada ≥ 10 mm
boyutundaki folliküllerin LH‘ye yanıt vermeye baĢladığını belirtmiĢlerdir.
Ayrıca çalıĢmada anovulatuvar polikistik ovaryumlu hastalarda granuloza
hücrelerinin LH‘ye normalden daha önce yanıt verdiklerini saptamıĢlar ve
bu erken yanıtın anovulasyon düzeneğinde rolü olduğunu bildirmiĢlerdir.
133
Filicori ve arkadaĢlarının 2002‘de yaptıkları çalıĢmada;
ovulasyon indüksiyonunun son evrelerindeki düĢük doz hCG Ģeklindeki LH
erkinin ovaryum follikülünün iĢlevini düzenlediği ve büyümesini sağladığı
varsayımını araĢtırmıĢlardır. Bunun için hastalara 7 gün süresince birinci
gruba sadece r-FSH 150 IU/gün; ikinci gruba r-hFSH 50 IU/gün ve hCG 50
IU/gün; üçüncü gruba, r-hFSH 25 IU/gün ve hCG 100 IU/gün; dördüncü
gruba ise sadece hCG 200 IU/gün uygulanmıĢtır. Sonuç olarak FSH‘nin
kesilip LH‘nin kullanılmasıyla follikülogenezisin uyarıldığı, folliküllerin
geliĢmeyi sürdürdüğü, küçük folliküllerin (<10 mm) sayısının azaldığı
gözlenmiĢtir. 134
Klomifen sitrat (KS) ilk olarak 1956 yılında sentezlenmiĢtir.
Klinik denemeleri ilk olarak 1960‘ da baĢlatılmıĢ ve ABD‘de klinik kullanımı
1967 yılında onaylanmıĢtır. KS kimyasal olarak, bir nonsteroidal
trifeniletilen yapısında olup, östrojen eĢdeĢi ve karĢıtı etkileri birlikte
barındırır. Klomifen sitrat iki farklı steroizomerin karıĢımıdır, farmakolojik
olarak cis ve trans yapıları vardır; enklomifen (cis-klomifen) ve zuklomifen
(trans-klomifen) olarak adlandırılır. Enklomifen en etkili izomerdir ve
ovulasyonu sağlayan etkiyle yükümlüdür. Enklomifenin yarı ömrü kısadır
ve serum yoğunluğu tedaviden sonra hızlıca yükselir ve birkaç gün sonra
125
düĢer. Zuklomifen daha yavaĢ atılır. Tek dozdan sonra bile haftalarca
serumda saptanabilir. ArdıĢık tedavilerde ovaryumda birikebilir. 51,52,53,54
Klomifen
sitrat
tedavisi
anovulatuvar
infertilitesi
olan
kadınlarda tedavide ilk yeğlenen ilaçtır. Anovulatuvar infertilitede KS
tedavisi sadece dolaĢan östrojen düzeyi gonadotropinler üzerinde negatif
geri bildirim oluĢturacak düzeye geldiğinde etkilidir. Klomifen sitrat ile
yapılan ovulasyon indüksiyonlarında memnun edici ovulasyon oranlarının (
%60-90) elde edildiği ancak gebelik oranlarının düĢük olduğu ( % 10-40)
bilinmektedir. 55,135
Imani ve arkadaĢları 201 oligomenore ya da amenoreli hasta
ile yaptıkları çalıĢmalarında 50mg/gün, 100mg /gün ve 150 mg/gün artan
dozlarda 3 menstrüel döngü izlemiĢler ve 156 hastada( %78) ovulasyon
elde etmiĢlerdir. 135
Fatemi ve
arkadaĢlarının
klomifen
sitrat
ve
letrozolü
karĢılaĢtırdıkları çalıĢmalarında, hastalara menstrüel döngülerinin 3-7
günleri arasında 2.5 mg/gün letrozol ya da 100 mg/gün klomifen sitrat
uygulamıĢlardır. Letrozol grubunda LH pikinin olduğu gün progesteron ve
östradol
düzeylerinin
klomifen
grubuna
karĢın
düĢük
olduğunu
belirtmiĢlerdir. Belirgin olarak klomifen sitratın follikül geliĢmesini daha iyi
uyardığını saptamıĢlardır. 136
Klomifen sitrat ve rFSH ile ovulasyon indüksiyonu uygulanan
tedavilerin sonuçlarına bakıldığında anovulatuvar kadınlarda gebelik oranı
%15 kadardır.
137
126
Bayram ve arkadaĢları klomifen dirençli polikistik over
sendromlu hastalarda ovulasyon, gebelik, çoklu gebelik ve OHSS
açısından
rekombinant
FSH
ve
üriner
FSH‘ı
karĢılaĢtırdıkları
çalıĢmalarında iki grup arasında anlamlı bir fark bulamamıĢlardır.
138
Biz de çalıĢmamızda oluĢturduğumuz gruplarda yaptığımız
istatistiksel değerlendirmede, primordiyal ve sekonder follikül sayılarında,
hormon uygulaması yapılan grup ve hormon uygulaması yapılan gruba ait
gebe grubu ile kontrol grubu açısından anlamlı bir artıĢ olmadığını
belirledik. Klomifen sitrat uygulaması yapılan grup ile klomifen sitrat
uygulaması yapılan gruba ait kontrol grubu karĢılaĢtırmasında da klomifen
sitrat uygulamasının primordiyal follikül ve sekonder follikül sayılarında
istatistiksel olarak anlamlı bir fark olmadığını belirledik. Klomifen sitrat
uygulaması yapılan gruba ait gebe grubunda ise kontrol grubuna karĢın bir
artıĢ olduğunu ve bu farkın klomifen sitrat uygulaması yapılan gruba karĢın
daha yüksek olduğunu gördük.
ÇalıĢmamızda ovaryum çaplarını karĢılaĢtırmak amacıyla
yaptığımız istatistiksel değerlendirmede, hormon uygulaması yapılan grup
ve hormon uygulaması yapılan gruba ait gebe grubunda hormon kontrol
grubuna karĢın anlamlı bir artıĢ olduğunu belirledik. Klomifen sitrat
uygulanan grup ve klomifen sitrat uygulanan gruba ait gebe grubunda
ovaryum çapları değerlendirildiğinde ise klomifen sitrat kontrol grubuna
karĢın anlamlı bir artıĢ göstermediğini izledik.
127
Tüm bu bulgulara ek olgun (Graaf) follikül çaplarını
karĢılaĢtırmak amacıyla yaptığımız istatistiksel değerlendirmede ise
hormon uygulaması yapılan grupta hormon kontrol grubuna karĢın bir artıĢ
gördük. Hormon uygulaması yapılan gruba ait gebe grubunda ise anlamlı
bir artıĢ olmamakla birlikte klomifen sitrat uygulaması yapılan grup ve
klomifen sitrat uygulaması yapılan gruba ait gebe grubunda da Graaf
follikül çaplarında istatistiksel olarak bir artıĢ belirlenmiĢtir.
PCNA, 36 kDa ağırlığında türler arasında iyi korunmuĢ,
hücre döngüsü için esas olan düzenleyicidir. Bugüne değin yapılan
çalıĢmalarda DNA sentezi sırasında DNA hasarlarının onarılması ve S
fazında DNA polimeraz delta için ko-faktör olarak iĢlev gördüğü
gösterilmiĢtir. 61,62,63
PCNA ekspresyonu, hücre döngüsünün G1 evresinde baĢlar,
en yüksek düzeye S fazında ulaĢır ve G2/M fazında da azalır. Dinlenme
evresindeki hücreler farklı mitojenlerle uyarıldığında aynı derecede PCNA
ve DNA sentezi yaptıkları gösterilmiĢtir. Biyolojik yarı ömrü yaklaĢık 20
saattir. Aldehitleri kullanarak fikse edilmiĢ hücrelerde ise PCNA‘nın
dağılımı farklıdır ve hücre döngüsü süresince yoğun yaygın çekirdek
boyanması
saptanır.
Ovaryumda
mayotik
bölünme
halinde
olan
hücrelerde, çekirdek ve sitoplazmada belirlenmiĢtir. Gonadotropinler ve
östradiol
granüloza
hücrelerinin
çoğalma
evresini
baĢlatabilirler.
61,62,64,65,66,67
Tomanek ve arkadaĢları domuzlarda yaptıkları çalıĢmada
domuz
primordiyal
folliküllerinin
oositlerinde
PCNA
salınımına
rastlamamıĢlardır. Ancak primer, sekonder ve erken antral folliküllerde
128
PCNA salınımı farklılığı belirlemiĢlerdir. Bunu da primordiyal folliküllerin
oositlerinin hareketsiz olmasına ve henüz genomik DNA eĢleĢmesi
göstermemelerine bağlamıĢlardır. 63
Feranil ve arkadaĢları buffalo ve kedi ovaryumlarında
yaptıkları çalıĢmada sağlıklı folliküllerin PCNA tutulumunun erken ve geç
atretik folliküllerin tutulumundan daha fazla olduğunu belirlemiĢlerdir.
Ayrıca atrezi süresince teka hücrelerinde PCNA tutulumunun oldukça
azaldığını göstermiĢlerdir. 139
Oktay
ve
arkadaĢları
yaptıkları
immunohistokimyasal
çalıĢmada sıçan ovaryumunda PCNA ve follikül büyümesi arasındaki
iliĢkiyi araĢtırmıĢlardır. Primordiyal folliküllerde oositte ve granüloza
hücrelerinde
immünreaktivite
izlememiĢlerdir.
Primer
follikülden
baĢlayarak, preantral ve antral folliküllerde granüloza hücrelerinde, oositte
ve teka hücrelerinde boyanma görülmüĢtür. Follikül büyüdükçe PCNA
boyanmasının da arttığını izlemiĢlerdir. PCNA‗nın hücre döngüsünü
düzenleyici etkisi olduğunu ve follikül büyümesinde önemli bir belirteç
olduğunu belirtmiĢlerdir. 76
Picut
ve
immunohistokimyasal
arkadaĢları,
çalıĢmada,
sıçan
ovaryumlarında
primordiyal,
primer,
yaptıkları
sekonder
ve
preovulatuvar follikülerde PCNA immunoreaktivitesi gözlemlemiĢlerdir.
Follikül sayımı için H&E boyanması ve PCNA karĢılaĢtırması yapmıĢlardır.
PCNA ‗nın follikülleri belirlediği için sayımı kolaylaĢtırarak zaman kazancı
sağladığını bildirmiĢlerdir. 140
129
Muskhelishvili ve arkadaĢları, Picut‘a benzer Ģekilde sıçan
ovaryumunda
follikül
sayımını
kolaylaĢtırmak
immunohistokimyasal boyama denemiĢlerdir.
için
PCNA
ile
Sonuç olarak primer,
sekonder ve antral folliküllerin oositlerinde ve granuloza hücrelerinde
immunboyanma olduğunu saptamıĢlardır. Ayrıca primordiyal folliküllerin
oositlerinde de zayıfta olsa boyanma olduğunu gözlemlemiĢlerdir. Bu
bulgunun, PCNA‘nın sadece proliferasyonda değil aynı zamanda hücre
DNA hasar tamiri düzeneğinde de rol oynadığının bir göstergesi olduğunu
bildirmiĢlerdir. 75
Bizim çalıĢmamızda da bu çalıĢmalara koĢut olarak, tüm
gruplarda PCNA immünreaktivitesi değerlendirilmiĢtir. Hormon uygulaması
yapılan gruplara ait kontrol ovaryumlarında, primordiyal folliküllerde oosit
çekirdek
ve
çekirdekçiğinde,
follikül
hücre
çekirdeklerinde
PCNA
tutulumunun belirgindi ve orta dereceli olduğu görülmüĢtür. Atretik
folliküllerde, granüloza hücrelerinde çekirdek tutulumu görülmekle birlikte
yine teka interna hücrelerindeki tepkime belirgin olarak izlenmiĢtir. Büyük
büyültmeli resimlerde primordiyal, tek sıralı hücre katmanı içeren primer ve
çok sıralı hücre katmanı içeren primer folliküllerde oosit çekirdeği,
granüloza hücre çekirdeği ve teka interna hücre çekirdeklerindeki tutulum
belirgin olarak gözlemlenmiĢtir. Antrumu belirgin sekonder folliküllerde
oositteki PCNA tutulumunun belirgin olarak azaldığı, granüloza hücre
tutulumu ile teka interna hücre tutulumunun ise oldukça yoğun olduğu ayırt
edilmiĢtir. Bazı stromal hücrelerde de zayıftan ortaya değiĢen tutulum
görülmüĢtür. Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryumlarında da PCNA tutulumunun hormon uygulaması yapılan
gruplara ait kontrol grubu ovaryumlarına eĢdeĢ olduğu görülmüĢtür.
Hormon uygulaması yapılan gruplara ait ovaryum dokusunda PCNA
immünreaktivitesinin, geliĢen folliküllerde oosit çekirdeği düzeyinde
kuvvetli olduğu, buna karĢın sekonder follikülden itibaren oosit çekirdek
130
boyanmasının azaldığı belirlenmiĢtir. Tüm folliküllerde granüloza ve teka
interna hücrelerinde yaygın tutulum dikkati çekerken, tutulumun çekirdek
düzeyinde fazla sitoplazmik düzeyde ise daha az olduğu izlenmiĢtir.
Klomifen sitrat uygulaması yapılan gruba ait ovaryum dokularında PCNA
tutulumunun tüm folliküllerde, atretik olanlarda stromada ve korpus
luteumda son derece az düzeyde olduğu dikkati çekmiĢtir. Granüloza
hücrelerinde ise bu tepkime çekirdek düzeyinde ve az yoğun olarak
belirlenmiĢtir.
Çok
sıralı
primer
folliküllerde
birden
fazla
oositin
gözlemlenmesi dikkati çekmiĢtir. Hormon uygulaması yapılan gruba ait
gebe ovaryumlarındaki PCNA tutulumu, tüm folliküllerde granüloza
hücreleri, teka interna hücreleri düzeyinde ve oosit çekirdeklerinde oldukça
belirgindi.
Ġmmünreaktivite
korpus
luteum
hücrelerinde
ve
atretik
folliküllerdeki hücrelerde yaygındı. Bazı stromal hücrelerde de oldukça
kuvvetli PCNA tutulumu dikkat çekiciydi. Klomifen sitrat uygulaması
yapılan gruba ait gebe ovaryumlarında ise PCNA immünreaktivitesinin,
tüm folliküllerde granüloza ve teka interna hücreleri düzeyinde olduğu ayırt
edilmiĢtir.
Oosit
Ġmmünreaktivitenin
çekirdek
korpus
ve
sitoplazmasında
luteumda
daha
az
tutulum
belirgindi.
düzeyde
olduğu
belirlenmiĢtir. GeliĢen folliküllerde iki oositin varlığı ayırt edilmiĢtir.
c-fos geninin transkripsiyonunu ilk kez 1984 yılında Mike E.
Greenberg ve Ed B. Ziff büyüme faktörleri ile uyardıkları fibroblastlarda
göstermiĢlerdir.
C-fos geninin transkripsiyonu hücreleri etkileyerek
büyüme ve farklılaĢmayı sağlayan çok sayıda etken tarafından hızla ve
geçici olarak sağlanır. C-fos bu nedenle hücrede herhangi bir uyarana
karĢı ilk yanıt veren genlerdendir [erken–ivedi geni, (IEG) ya da erken
yanıt geni, (ERG)]. Bu gruba giren genler hücre büyümesi, çoğalması,
farklılaĢması
ve
hücrenin
programlı
ölümü
(apopitozis)
ile
ilgili
transkripsiyon faktörleridir. 77,78,79,80
131
C-fos transkripsiyonu çok çeĢitli mitojenler ve farklılaĢmayı
uyaran maddeler tarafından uyarılabilir ve bağımsız olarak yeni protein
sentezi oluĢturur. Bu nedenle bu tür genler protoonkojenler ve bunların
ürünü olan proteinler de onkoproteinler olarak adlandırılırlar. Kanser
geliĢiminde Fos proteinlerinin katkısı olduğu bilinmektedir. Bazı tümör
tiplerinde c-fos ve c-jun düzeyinin arttığı gösterilmiĢ ve bu artıĢın artan
hücre çoğalmasının bir kanıtı olacağı için tümör oluĢmasında önemli bir rol
oynadığı ileri sürülmüĢtür. Ancak protoonkojenler yalnız kötü huylu geliĢim
ile birlikte değil, normal büyüme ve farklılaĢmayı da düzenlerler.
91, 98,99
Mahner ve arkadaĢlarının ovaryum karsinoma hücreleriyle
yaptıkları çalıĢmada c-fos immunboyanması tümör hücre çekirdeklerinde
ve bazı stromal fibroblastların çekirdeklerinde saptanmıĢtır.
141
AP-1 kompleksin üyeleri değiĢik gen ekspresyonlarını
düzenlerler. Bu bileĢenler çoğalma, farklılaĢma, programlı hücre ölümü ve
onkojenik transformasyon gibi çok sayıda hücresel süreçlerle kritik olarak
ilgilidir.
Tarlov ve arkadaĢları AP-1 ailesinin özellikle c-fos ve c-jun
genlerinin hücrede apoptozisi düzenledikleri varsayımını araĢtırmıĢlardır.
C-fos ve c-jun genlerine mutasyon yaptıkları fare embriyolarında in situ
apopitozis deneyi uygulamıĢlardır. Sonuç olarak bu genlerin yokluğunda
da
programlanmıĢ
hücre
ölümünün
embriyolarda
gerçekleĢtiğini
bildirmiĢlerdir.142
Fos
proteinleri
c-fos
geninin
ürünü
olan
çekirdek
proteinleridir. Bunların hücredeki ekspresyonu ve birikimi herhangi bir etki
132
olmaksızın görülebilir ancak bu ekspresyon ve birikim çeĢitli uyarılara yanıt
olarak hücre erkiyle birlikte artar.
Sharma ve arkadaĢlarının yaptığı çalıĢmada; hipofizektomi
yapılmıĢ sıçanlara follikül büyümesini ve luteinizasyonunu uyarmak için
FSH ve LH uygulanmıĢtır. AP1 faktörleri için immunohistokimyasal
uygulama yapılmıĢtır. Jun (c-Jun, JunD ve JunB) ve Fos ailesi (c-Fos,
FosB, Fra1ve Fra2) üyelerinin granuloza hücrelerinde, teka hücrelerinde
ve luteal hücrelerde varlığı belirlenmiĢtir.
143
Bizim çalıĢmamızda ise hormon uygulaması yapılan gruba
ait
kontrol
grubu
ovaryumlarında
c-FOS
immünreaktivitesi
değerlendirildiğinde, tutulumun tüm folliküllerde oosit sitoplazması ile
çekirdeğinde ve granüloza hücreleri ile teka interna hücrelerinde olduğu
görüldü. Tutulumun çekirdek ve sitoplazma düzeyinde granüler Ģekilde
olduğu ve çoğunlukla orta dereceli gözlemlendiği ilgiyi çekti. Bu gruba ait
incelemelerde korpus luteum hücrelerinde de orta dereceli immünreaktivite
belirgindi. Klomifen sitrat uygulaması yapılan gruba ait kontrol grubu
ovaryumlarındaki c-fos immünreaktivitesi de, hormon grubu kontrolleri ile
eĢdeĢ yapı sergiliyordu. Hormon uygulaması yapılan gruplara ait
ovaryumlarda c-fos immünreaktivitesi değerlendirildiğinde, tutulumun
kontrole karĢın biraz daha çok olduğu ve çekirdek ile sitoplazma
düzeyinde yaygın olduğu belirlendi. Hormon uygulaması yapılan gruplara
ait gebe ovaryumlarında ise c-fos tutulumunun diğer gruplara karĢın biraz
daha özgün olduğu, sitoplazma ve çekirdekte granüller halinde belirdiği
ilgiyi çekti. Klomifen sitrat uygulaması yapılan grup ve hormon uygulaması
yapılan grup karĢılaĢtırıldığında, klomifen sitrat uygulması yapılan grupta
c-fos tutulumunun ağırlıklı olarak teka interna hücreleri ve stromada
yoğunlaĢtığı izlendi. Tutulumun bu grupta tüm folliküllerde, granüloza
133
hücreleri ve oositte, hormon grubuna karĢın az olması ilgiyi çekerken,
korpus luteum hücrelerinde biraz daha yoğunlaĢması dikkati çekti.
Klomifen sitrat uygulaması yapılan gruba ait gebe grubu ovaryum
dokularında çok sıralı antrumun oluĢmaya baĢladığı, folliküllerde c-fos
tutulumunun belirgin olarak arttığı ve granüloza hücrelerinde, oosit ile teka
interna hücrelerinde yaygınlaĢtığı belirlendi. Tutulum sitoplazmada ve
granüler Ģekildeydi.
Lösemi inhibitör faktör (LIF) 43 kDa moleküler ağırlığa sahip
bir glikoproteindir. Biyolojik erki çeĢitli dokularda bulunur. LIF, embriyonik
kök hücrelerin, primordiyal germ hücrelerinin, periferal nöronların,
osteoblastların, adipositlerin, hepatositlerin ve endotelyal hücrelerin
büyümesini ve farklılaĢmasını düzenler. LIF, fibroblastlar, alloreaktif Tlenfositler, mitojen erkli dalak hücreleri, Krebs ve Ehrlich asit mayi
hücrelerince üretilir. Hücre aktivasyonundan sonra monositlerce de üretilir.
LIF‘in mezenĢimal hücrelerde sentezi, IL-1α, TGF-β, EGF ve bFGF
tarafından
uyarılır.
Folliküler
sıvıdaki
granuloza-lutein
hücreleri,
makrofajlar ve monositler insan folliküler hücrelerinin % 5-15‘ini oluĢturur
ve bu hücreler LIF kaynağı olarak bilinmektedirler. Ayrıca bir diğer
potansiyel LIF kaynağı da follikülü çevreleyen teka hücreleridir. 108,109
Yapılan in vitro çalıĢmalarda LIF‘in fare blastosistlerinin
geliĢiminde rol oynadığı ortaya konulmuĢtur. Kemirgenlerde yapılan
çalıĢmada erken embriyo kültürlerinde LIF‘in trofoektoderm kütlesinin
artmasına neden olduğu gösterilmiĢtir. 144,145
134
Sığır embriyo deneylerinde, in vitro kültür ortamına insan LIF
proteini eklenmiĢ, sonuç olarak blastomerlerin sayısının arttığı, embriyo
dejenerasyon yüzdesinin düĢtüğü ve implantasyon yüzdesinin ve gebelik
oranının arttığı izlenmiĢtir. Transgenik farelerde yapılan bir diğer
çalıĢmada, LIF yokluğunda farelerde normal embriyoların geliĢebildiği
ancak implante olamadıkları belirlenmiĢtir.
146
Abir ve arkadaĢları yaptıkları çalıĢmada LIF‘in büyüme
faktörü olarak ovaryumdaki primordiyal folliküllere etkisi olup olmadığını
araĢtırmıĢlardır. Gruplarını 19-33 haftalık düĢük insan fetusu ve 13-43
yaĢları arası premenapozal hastalardan oluĢturmuĢlardır. LIF ve LIF
reseptörleri olan LIF-R ve gp 130‗u immunohistokimyasal olarak
belirlemeye çalıĢmıĢlardır. Sonuçta her iki grupta da primordiyal, primer ve
sekonder folliküllerin oositlerinde zayıf gp-130 tutulmasına karĢın güçlü
LIF ve LIF-R tutulması saptamıĢlardır. YetiĢkin ovaryumlarında primer ve
sekonder folliküllerin granuloza hücrelerinde LIF-R‘yi belirlemelerine karĢın
fetuste LIF-R‘ye rastlamamıĢlardır. Her iki grupta da bazı stromal
hücrelerde LIF tutulumu tespit edilmiĢtir. AraĢtırıcılar LIF ve reseptörlerinin
yetiĢkin ve fetal primordiyal folliküllerin oositlerinde bulunmasının, LIF‘in
diğer
büyüme
faktörleri
ile
düĢündürttüğünü belirtmiĢlerdir.
birlikte
follikülogenezisi
ilerlettiğini
147
Arıcı ve arkadaĢları, ÜYTE tedavisi gören ve embriyo
transferi gerçekleĢtirilen hastalardan alınan folliküler sıvı örnekleri
kullanarak LIF, östradiol ve progesteron değerlerini ölçmüĢ ve embriyo
kalitesini incelemiĢlerdir. ÇalıĢma sonucunda LIF mRNA ve protein
düzeyleri değerlendirilmiĢtir. Ovaryum stromal hücre kültürlerinde zayıfta
olsa LIF mRNA ve proteininin eksprese olduğu izlenmiĢtir. Aynı zamanda
bu ekspresyonun IL-1α ve TNF-α varlığında yükseldiğini de saptamıĢlardır.
135
ÇalıĢmada saflaĢtırılmıĢ granuloza hücrelerinin de düĢük miktarda LIF
mRNA ve proteini eksprese ettiği bildirilmiĢtir. AraĢtırıcılara göre LIF‘in bu
hücrelerde bulunması ovulasyon fizyolojisi, östrojen üretimi ve erken
embriyo geliĢiminde rol oynadığını göstermektedir.
148
Bizim çalıĢmamızda klomifen sitrat uygulaması yapılan gruba
ait kontrol grubuna ait ovaryum dokusu incelemelerinde LIF tutulumunun
hormon uygulaması yapılan grubun kontrol grubuyla eĢdeĢ olduğu
görüldü. Hormon uygulaması yapılan gruba ait ovaryum dokusunda
yapılan incelemelerde de LIF immünreaktivitesinin, kontrollere eĢdeĢ
olduğu dikkati çekiyordu. Hormon uygulaması yapılan gruba ait gebe
grubu ovaryum dokularında, bazı folliküllerde granüloza hücreleri, teka
interna hücreleri ve atretik follikül hücrelerinde diğer gruplara karĢın
tutulumun biraz daha yaygın olduğu belirlendi. Klomifen sitrat uygulaması
yapılan
gruba
ait
ovaryumlardaki
değerlendirmelerde,
LIF
immünreaktivitesinin korpus luteum hücrelerinde biraz daha çok olduğu
dikkati çekerken, diğer yapıların kontrol grubundakilerle aynı görünümde
olduğu belirlendi. Sekonder folliküllerde ise oositi çevreleyen ilk sıra
hücrelerde
tutulumun
biraz
daha
arttığı
görüldü.
Klomifen
sitrat
uygulaması yapılan gruba ait gebe gruplarında yapılan incelemelerde ise,
LIF immünreaktivitesinin diğer gruplara karĢın artmıĢ olması ilgiyi çekti.
Tutulum oosit sitoplazmasında az, granüloza hücrelerinde ve teka interna
hücrelerinde ise biraz daha fazlaydı.
136
6. SONUÇ
Klomifen sitrat ile gonodotropinlerin ovulasyon induksiyonu
için kullanımları sonucunda ovaryum dokusunda neden olabilecekleri
değiĢikliklerin incelendiği çalıĢmamızda, primordiyal ve sekonder follikül
sayısının, hormon uygulaması yapılan grup, bu grubun kontrol ve
gebelerinde istatistiksel olarak anlamlı bir fark göstermediği saptanmıĢtır
(p=0.354, p=0.795). Primordiyal ve sekonder follikül sayısının klomifen
sitrat uygulaması yapılan grup ve klomifen sitrat uygulaması yapılan gruba
ait kontrol grubunda aynı olduğu belirlenmekle birlikte, klomifen sitrat
uygulaması yapılan gruba ait gebe grubunda istatistiksel olarak arttığı
belirlenmiĢtir (p=0.802, p=0.582). Bu bulgulara koĢut olarak primordiyal ve
sekonder follikül sayısı değerlendirildiğinde; hormon ve klomifen sitrat
uygulamalarının bu folliküllerin sayısında bir değiĢikliğe neden olmadığı,
ancak klomifen sitrat uygulamasını izleyerek gebe kalan grupta follikül
sayısının arttığı istatistiksel olarak saptanmıĢtır.
Ovaryum çaplarının gruplar arası istatistiksel değerlendirmesi
sonucunda; hormon uygulaması yapılan grup ve hormon uygulaması
yapılan gruba ait gebelerde, hormon uygulaması yapılan gruba ait kontrol
grubuna karĢın anlamlı bir artıĢ gösterdiği saptanmıĢtır (p=0.180) Klomifen
sitrat uygulaması yapılan ve klomifen sitrat uygulaması yapılan gruba ait
gebelerde ise ovaryum çaplarının, klomifen sitrat uygulaması yapılan
gruba ait kontrol grubu ile aynı olduğu belirlenmiĢtir (p=0,565). Sonuç
olarak klomifen sitrat uygulamasının, gebelik de dahil olmak üzere
ovaryum çapında değiĢiklik yapmadığı, buna karĢın hormon uygulaması
yapılan grup ve hormon uygulaması yapılan gruba ait gebe grubunda, bu
grubun kontrol grubuna karĢın anlamlı artıĢ gösterdiği istatistiksel olarak
belirlenmiĢtir.
137
Graaf follikülü çaplarının istatistiksel değerlendirmesinde;
hormon uygulaması yapılan grupta, hormon uygulaması yapılan gruba ait
kontrol grubuna karĢın artıĢ olduğu (p=0.095), gebe grubunun ise kontrol
grubuna yakın olduğu belirlenmiĢtir. Hem klomifen sitrat uygulaması
yapılan grup hem de klomifen sitrat uygulaması yapılan gruba ait gebe
grubundaki fark ise istatistiksel olarak anlamlı bulunmuĢtur (p=0.006).
Sonuç olarak hormon uygulamasının Graaf follikül çapını artırdığı,
gebelikte ise bir değiĢikliğe neden olmadığı belirlenirken, klomifen sitratın
hem klomifen sitrat uygulaması yapılan grupta hem de klomifen sitrat
uygulaması yapılan gruba ait gebelerde Graaf follikül çapını artırdığı
istatistiksel olarak saptanmıĢtır.
ÇalıĢmamızda PCNA immünreaktivitesinin değerlendirilmesi
sonucunda; hormon uygulaması yapılan gruba ait kontrol ovaryumlarında
primordiyal folliküllerde oosit çekirdeğinde, follikül hücre çekirdeklerinde ve
teka interna hücre çekirdeklerinde boyanmanın orta dereceli olduğu
gözlemlenmiĢtir. Atretik follüküllerde de tüm granüloza hücrelerinde
çekirdek tutulumu izlenmiĢtir. Antrumu belirgin sekonder folliküllerde de
oldukça yoğun tutulum ayırt edilmiĢtir. Stromal hücre immün boyaması da
dikkati çekmiĢtir. Yine bu grupta PCNA tutulumunun antrumu belirgin
sekonder folliküllerden itibaren, oosit çekirdeğinde azaldığı ancak follikül
ve teka interna hücrelerinde yaygın olduğu ayırt edilmiĢtir. Klomifen sitrat
uygulanan grupta ise tutulumun tüm yapılarda daha az olduğu
gözlemlenmiĢtir. Sonuç olarak PCNA‘nin ovaryum folliküllerinde sayım için
iyi bir belirteç olduğu, primer, sekonder ve antral folliküllerde belirgin
boyanma gösterdiği belirlenmiĢtir. Yine tüm oositlerin follikül geliĢiminde
zayıftan kuvvetliye değiĢen boyanma gösterdiği saptanmıĢ ve bunun
PCNA‘ nın yalnızca proliferasyonda değil, DNA hasar düzeneğinde de rol
oynadığının bir göstergesi olabileceği yargısına varılmıĢtır.
138
Normal ve patolojik hücre çoğalma belirteci olan c-fos‘un,
ovaryum indüksiyonunda proliferasyonun iĢaretleyicisi olarak kullanılması
sonucunda; hormon uygulaması yapılan grupta diğer gruplara karĢın daha
özgün tutulum gösterdiği gözlenmiĢtir. Özellikle hormon uygulanan gebe
ovaryumlarında korpus luteum‘da belirgin artıĢ izlenmiĢtir. Tutulumun
klomifen sitrat uygulaması yapılan gruplarda ise, hormon uygulaması
yapılan gruplara karĢın daha az olduğu belirlenmiĢtir. Klomifen sitrat
uygulaması yapılan gebe ovaryumlarında ise hormon uygulaması yapılan
gebe gruba yakın dağılım gösterdiği saptanmıĢtır. Sonuç olarak hormon
uygulaması yapılan grupta c-fos tutulumunun belirgin artıĢı, hormonun
aĢırı hücre proliferasyonuna neden olduğunun ve c-fos‘un bu konuda
önemli bir belirteç olarak kullanılabileceğinin göstergesi olarak kabul
edilmiĢtir.
LIF dağılımının gruplara göre değerlendirilmesi sonucunda,
hormon uygulaması yapılan gruplardaki gebe sıçanların ovaryumlarında
belirgin artıĢ gösterdiği saptanırken, klomifen sitrat uygulaması yapılan
gruba ait kontrol grubuna karĢın, klomifen sitrat uygulaması yapılan grup
ve klomifen sitrat uygulaması yapılan gruba ait gebe grupta tutulumun
arttığı izlenmiĢtir. Sonuç olarak LIF‘in follikülogenezisin ilerletilmesinde ve
erken embriyonik dönemde etkin olduğu düĢünülmüĢ, yine östrojen
üretiminde de varlığının arttığı saptanmıĢtır.
Tüm bu bulgular yapısal olarak östrojene benzeyen klomifen
sitratın östrojen reseptörlerine bağlanarak reseptörleri kapattığı, östrojene
karĢı
duyarlılıkta
azalmaya
koĢut
olarak
da
reseptör
sayısını
azaltabileceğini düĢündürmüĢtür. Klomifen sitratın follikül geliĢiminden çok
oosit
geliĢiminde
etkili
olduğu
sonucuna
varılmıĢtır.
Hormonal
indüksiyonda ise ovaryumun daha çok hücresel düzeyde etkilendiği
düĢünülmüĢtür.
139
7. ÖZET
Ġnfertilite; belirli bir doğum kontrol yöntemi uygulamadan,
haftada en az iki kez düzenli cinsel yaĢama karĢın bir yıl süreyle gebelik
oluĢmaması olgusudur. Ġnfertilite sorunu üreme çağındaki çiftlerin yaklaĢık
% 10-15‘ini etkilemektedir. Kadınlardaki en önemli infertilite nedeni
yaklaĢık olarak % 40 oranında ovulasyon yetersizliğidir. Günümüzde
ovulasyon indüksiyonunda amaç, anovulatuar hastalarda ovulasyonu
baĢlatmaktır. Bu konuyla ilgili olarak yapılan çalıĢmalarda üst üste yapılan
hormonal ovulasyon indüksiyonu uygulamalarının önlenemez hücre
çoğalmasına ve kansere neden olduğu da bildirilmiĢtir.
Bizim çalıĢmamızda klomifen sitrat ve gonodotropinler ile
ovulasyon indüksiyonu uygulanmıĢ ve geliĢebilecek histolojik değiĢimlerin
ve hasarın, ovaryum dokusunda hücre proliferasyon iĢaretleyicileri olan
PCNA, LIF ve c-FOS belirteçleri kullanılarak immunohistokimyasal olarak
karĢılaĢtırmalı incelenmesi amaçlanmıĢtır.
Yapılan immünohistokimyasal değerlendirmelerde; klomifen
sitrat ve gonadotropinlerin ovulasyon indüksiyonu için kullanımları
sonucunda ovaryum dokusunda, oosit düzeyinde yapısal değiĢiklikler ve
çoklu oosit oluĢumuna neden olduğu görülmüĢtür.
Tüm bulgular yapısal olarak östrojene benzeyen klomifen
sitratın östrojen reseptörlerine bağlanarak reseptörleri kapattığı, östrojene
karĢı
duyarlılıkta
azalmaya
koĢut
olarak
da
reseptör
sayısını
azaltabileceğini düĢündürmüĢtür. Klomifen sitratın follikül geliĢiminden çok
140
oosit
geliĢiminde
etkili
olduğu
sonucuna
varılmıĢtır.
Hormonal
indüksiyonda ise ovaryumun daha çok hücresel düzeyde etkilendiği
düĢünülmüĢtür.
141
8. SUMMARY
Failure to conceive after 1 year despite regular sexual life
with two intercourses a week in the absence of any method of
contraception is defined as infertility. According to reports, infertility affects
almost 10-15% of couples during their reproductive ages. The most
prevalent cause of infertility in women is ovulation disorders, accounting
for 40% of all cases. In cases where ovulation is achieved, the main
purpose is single follicle ovulation. Related studies have shown that
successive hormonal ovulation inductions result in unavoidable cell
proliferation and cancer.
The present study aims to investigate histological changes
and damage that may result from treatment with clomiphene citrate and
gonadotropins for ovulation induction using cell proliferation markers
PCNA, LIF and c-FOS in the ovary tissue using immunohistochemical
methods.
Immunohistochemical analysis of potential damages in the
ovary tissue that clomiphene citrate and gonodotropins may cause as a
result of their use for ovulation induction demonstrated structural changes
in oocyte levels and, particularly, multiple oocytes.
All findings suggested that clomiphene citrate, which is
structurally similar to estrogen, closes estrogen receptors by binding to
them and this may result in a decreased number of receptors in parallel to
the decreased sensitivity to estrogen. In conclusion, we are in the opinion
142
that clomiphene citrate is effective on oocyte development rather than
follicle development.
143
9. KAYNAKLAR
1. Mosher WD, Pratt WF. Fecundity and infertility in the United
States:incidence and trends. Fertil.Steril 1991;56:192-93.
2. Berek JS. Novak‘s Gynecology. 12th press. Williams & Wilkins,
Baltimore, Maryland USA 1998; 27: 915-962.
3. Hughes E, Collins J, Vandekerckhove P. Clomiphene citrate for
ovulation induction in women with oligo-amenorrhoea. Cochrane
Dtabase Syst Rev 2000; 2:CD000056.
4. Scott RT, Jr., Hofmann GE. Prognostic assessment of ovarian reserve.
Fertil Steril 1995;63:1-11.
5. Hull MGR, Glazner CMA, Kelly NJ. Population study of causes,
treatment and outcome of infertility. Br Med J 1983; 291: 1693-1697.
6. Speroff L, Glass RH, Kase NG. Clinical Gynecologic Endocrinology and
Infertility. Lippincott Williams & Wilkins, Baltimore, Maryland, U.S.A
1999;6: 487-522.
7. Speroff L, Glass RH, Kase NG. Clinical Gynecology Endocrinology and
Infertility. Lippincott Williams & Wilkins,1999;6:1013-1132.
8. Aboulghar MA, Amin Y, Mansour RT, Abbas AM, Serour GI, Salah IM.:
Ovarian superstimulation and intrauterine insemiantion for the
tretament of unexplained infertility. Fertil Steril 1993;60:303-306.
9. Dickey RP, Olar TT, Taylor SN, Curole DN, Rye PH. Sequential
clomiphene citrate and human menopausal gonadotropin for ovulation
induction: comparion to clomiphene citrate alone and human
menopausal gonadotropin alone. Hum Reprod 1993; 8:56-59.
10. Agarwal SK, Buyalos RP. Clomiphene citrate with intrauterine
insemination: Is it effective therapy in women above the age of 35
years? Fertil Steril 1996; 65:759-763.
144
11. Kistner RW. Sequential use of clomiphene citrate and human
menopausal gonadotropin in ovulation induction. Fertil Steril 1976;
27:72-82.
12. Rao MC, Midgley AR, Richards JS. Hormonal regulation of ovarian
cellular proliferation 1978; 14: 71-78.
13. Kistner RW. Ovulation: clinical aspects. In Balin H. And Glasser S.
(eds) Reproductive Biology. Excerpta Medica, Amsterdam 1972:477
14. March CM, Tredway DR, Mishell DR. Effect of pretreatment with
clomiphene citrate upon human menopausal gonadotropin therapy for
anovulation. Fertil Steril 1975; 26:191-192.
15. Homburg R, Insler V. Ovulation induction in perspective. Hum Reprod
Update 2002; 8: 449-62.
16. Ohba T., Ujioka T, Ishikawa K, Nobuyuki T, Okamura H. Ovarian
hyperstimulation syndrome-model rats; the manifestation and clinical
implication. Molecular and Cellular Endocrinology 2003; 202: 47-52.
17. A. Corakcı, S. Fılız, E. Calıskan, C. Dalcık, S. Ozeren, H. Dalcık: The
effects of ovulation induction on ovarian epithelium dysplasia scores
and Ki67 expression: an experimental study on rats, Int J Gynecol
Cancer 2005, 15, 866–871.
18. Sadler T. Medikal Embriyoloji ( Türkçe Çeviri) 9. Baskı. Palme
Yayıncılık. Ankara.2005.
19. 32 Moore KL, Persaud TVN. Klinik Yönleri ile Ġnsan Embriyolojisi (
Türkçe Çeviri) 6. Baskı. Nobel Tıp Kitabevleri. Ankara. 2002.
20. Motta PM, Makabe S, Nottola SA. The ultrastructure of human
reproduction I. The natural history of the female germ cell : origin,
migration and differentiation inside the developing ovary. Hum.
Reprod Update 1997; 3: 281- 95.
21. Block E. Quantitative morphological investigations of the follicular
system in woman ; variations at different ages. Acta Anat ( Basel)
1952; 14: 108-23.
145
22. Kierszenbaum A L. Histoloji ve Hücre Biyolojisi ( Türkçe Çeviri). Palme
Yayıncılık. 2006.
23. Arıncı, Elhan A: Anatomi 1. Cilt. Ankara. GüneĢ Kitabevi Lmt; 2001.
24. Erdoğan D, Hatipoğlu MT, Görgün M, Ilgaz C. Özel Histoloji. Hatipoğlu
2007.
25. Ben Cherit A, Gotlieb L, Wong PY, Casper RF. Ovarian response to
recombinant
human
follicle-stimulating
hormone
in
luteinizing
hormone-depleted women: examination of two cell, two gonadotropin
theory. Fertil. Steril 1996;65:711-17.
26. Garner LP, Hiatt JL. Color Textbook of Histology. Saunders 2001.
27. Ross M H, Kaye G I , Pawlina W. Histology a Text and Atlas.
Lippincott Williams&Wilkins 2003.
28. Junqueıra, Carneıro J. Temel Histoloji ( Türkçe Çeviri ) Nobel 2006.
29. Guyton A C, Hall J E. Text Book of Medical Physiology, International
Edition, W.B. Saunders 2006.
30. Ganong W.F. Tıbbi Fizyoloji. Nobel Tıp Kitabevleri, 20. Baskı. 2002.
31. Augood C, Duckitt K, Templeton AA. Smoking and female infertility: a
sistematic review and meta-analysis. Hum. Reprod 1998;13: 15321539.
32. Gomel V UB, Yarali H. Investigation of the infertile couple. In
Reproductive Endocrinology and Infertility 1993:143-155.
33. Ventura SJ, Hamilton BE, Sutton PD. Revised birth and fertility rates
for the United states, 200 and 2001. Natl. Vital Stat. Rep. 2003;51:118.
34. Quagliarello J, Arny M. Inaccuracy of basal body temperature charts in
predicting urinary luteinizing hormone surges FertilSteril 1986;45:334337.
35. Greenblatt RB, Barfield WE, Jugck E.C. and Ray A.W. Induction of
ovulation with MRL/41. Preliminary report. JAMA 1961; 178: 101-4.
146
36. Gemzell CA., Diczfalusy E. and Tillinger G. Clinical effect of human
pituitary follicle-stimulating hormone (FSH). J Clin Endocrinol Metab
1958;18: 1333-48.
37. Gemzell CA. Treatment of infertility after partial hypophysectomy with
human pituitary gonadotrophins. Lancet 1964; i: 644-647.
38.Bettendorf
G.
Human
hypophyseal
gonadotrophins
in
hypophysectomized women. Int J Fertil 1963; 8: 799.
39. Knobil E. Neuroendocrine control of the menstruel cycle. Rec Prog
Horm Res.1980; 36: 53-88.
40. Filicori M, Cognigni GE, Pocognoli P, Tabarelli C, Ferlini F, Perri T,
Parmegiani L. Comparison of controlledovarian stimulation with
human menapausal gonadotropin or recombinant follicle-stimulating
hormone. Fertil. Steril. 2003;80:390-397.
41. Rao MC, Midgley AR, Richards JS. Hormonal regulation of ovarian
cellular proliferation. 1978; 14: 71-8.
42. Ludwig M, Doody KJ, Doody KM. Use of recombinant human chorionic
gonadotropin in ovulation induction. Fertil Steril 2003; 79: 1051-9.
43. Schoot DC, Harlin J, Shoham Z, Mannaerts BM, Lahlou N, Bouchard
P, Bennink HJ, Fauser BC. Recombinant human follicle-stimulating
hormone and ovarian response in gonadotropin-deficient women.
Hum Reprod 1994;9:1237-1242.
44. Gemzell C. Induction of ovulation with human gonadotropins. Recent
Progr Horm Res 1965; 21: 179-204.
45. Mazer C, Ravetz E. Am J Obstet Gynecol 1941; 41: 474-84.
46. Van Der MM, Hompes PG, de Boer JA, Schats R, Schoemaker J.
Cohort size rather than follicle-stümulating hormone threshold level
determines ovarian sensitivity in polycystic ovary syndrome. J Clin
Endocrinol. Metab 1998;83:423-426.
147
47. Homburg R, Levy T, Ben Rafael Z. A comparative prospective study of
conventional regimen with chronic low-dose administration of folliclestimulating hormon efor anovulation associted with polycystic ovary
sendrome. Fertil Steril. 1995;63:729-733.
48. Lu PY, Chen AL, Atkinson EJ, Lee SH, Erickson LD, Ory SJ. Minimal
stimulation
achieves
pregnancy
rates
comparable
to
human
menapausal gonadotropins in the treatment of infertility. Fertil Steril.
1996;65:583-587.
49. Reuter KL, Cohen S, Furey l, Baker S. Sonographic appearance of the
endometrium and avaries during cycles stimulated with human
menapausal gonadotropin. J Reprod Med 1996;41:509-514.
50. Silverberg KM, Olive DL, Burns WN, Johnson JV, Groff TR, Schenken
RS. Follicular size at the time of human chorionic gonadotropin
administration predicts ovulation otcome in human menapausal
gonadotropin-stimulated cycles. Fertil Steril. 1991;56:296-300.
51. Dickey RP, Holtkamp DE. Development, pharmacology abd clinical
experience with clomiphene citrate. Hum Reprod Update 1996;2:483506.
52. Clark JH, Markaverich BM. The agonistic-antagonistic properties of
clomiphene: a review. Pharmacol Ther 1981;15:467-519.
53. Çolgar U. Reprodüktif Endokrinoloji ve Ġnfertilite. Ġstanbul Medikal
Yayıncılık 1. Baskı 2006.
54. Young SL, Opsahl MS, Fritz MA. Serum concentrations of
enclomiphene and zuclomiphene across consecutive cycles of
clomiphene citrate therapy in anovulatory infertile women. Fertil steril.
1999;7:639-644.
55. Glazener CM, Coulson C, Lambert PA, Watt EM, Hinton RA, Kelly NG,
Hull MG. Clomiphene treatment for women ith unexplailned infertility:
placebo-controlled study of hormonal responses and conception rates.
Gynecol Endocrinol.1990;4:75-83.
148
56. Fisch B, Margara RA, Winston RM, Hillier SG. Cellular basis of luteal
steroidogenesis in the human ovary. J Endocrinol 1989;122:303-311.
57. Gysler M, March CM, Mishell DR, Jr., Bailey EJ. A decade‘s
experience with an individualized clomiphene treatment regimen
including its effect on the postcoital test. Fertil. Steril. 1982;37:161167.
58. Imani B, Eijkemans MJ, te Velde ER, Habberna JD, Fauser BC.
Predictiors of chances to conceive in ovulatory patients during
clomiphene citrate induction of ovulation in normogonadotropic
oligoamenorrheic infertility. J Clin Endocrinol. Metab 1999;84:16171622.
59. Garcia J, Seegar Jones G, Wentz AC. The use of clomiphene citrate.
Fertil. Steril 1977;28:707-717.
60. Hsu CC, Kuo HC, Wang ST, Huang KE. Interference with uterine
blood flow by clomiphene citrate in women with unexplained infertility.
Obstet Gynecol 1995;866:917-921.
61. Tomanek M, Chronowska E. Immunohistochemical localization of
proliferating cell nuclear antigen (PCNA) in the pig ovary. Folia
Histochem Cyto. 2006, 44(4): 269-274.
62. Oktay K, Schenken RS, Nelson JF. Proliferating cell nuclear antigen
marks the initiation of follicular growth in the rat. Biol Reprod. 1995,
53: 295-301.
63. Tomanek M, Chronowska E. Immunohistochemical localization of
proliferating cell nuclear antigen(PCNA) in the pig ovary. Folia
Histochem Cytobol.2006;44: 269-274.
64. Bravo R, Macdonald Bravo H. Changes in the nuclear distribution of
cyclin (PCNA) but not its synthesis depend on DNA replication. EMBO
1985;4: 655-661.
65. Bravo R. Synthesis of nuclear protein cyclin and its relationship with
DNA replication. Exp Cell Res 1986;163:287-293.
149
66: Bravo R, Macdonald-Bravo H. Existance of two populations of
cyclin/proliferating cell nuclear antigen during the cell cycle.
Association with DNA replication sites. J Cell Bio 1987; 105: 15491554.
67. iyachi K, Friltzer MJ, Tan EM. Autoantibody to nuclear antigen in
proliferating cells. J Immunol 1978;121:2228-2234.
68. Ogata K, Ogata Y, Takasaki Y, Tan EM. Epitopes of proliferating cell
nuclear antigen recognized by human lupus autoantibody and murine
monoclonal antibody. J Immunol 1987;139:2942-2946.
69. Ogata N, Kamimura T, Asakura H. Point mutation, allelic loss and
increased methylation of c-Ha-ras gene in buman hepatocellular
carcinoma. Hepatology 1991;13:31-37.
70. Waseem NH, Lane DP. Monoclonal antibody analysis of the
proliferating cell nuclear antigen(PCNA).Structural conservation and
the detection of a nucleolar form. J cell Science 1990; 96: 121-129.
71. Dervan PA, Magee HM, Buckley C, Carney DN. Proliferating cell
nuclear antigen counts in formalin fixed parafin embedded tissue
corralate with Ki-67 in fresh tissue. Am J Clin Pathol 1992; 97:21-28.
72. Hall PA, Levision DA, Woods AL,Yu CC-W et al. Proliferating cell
nuclear antigen (PCNA) immunolocalization in parafin sections: An
index of cell proliferation with evidence of deregulated expression in
some neoplasms. J Pathol 1990; 62: 285-294.
73. Smith FG, Murray PG,Crocker J. Correlation between PCNA and
AgNOR scores in non-Hodkin‘s lymphomas using sequential staining
technique J Clin Pathol1993;46:28-31.
74.Woods AL, Hall PA, Shephard NA. The assesment of proliferating cell
nuclear antigen(PCNA) immunostaining in primary gastrointestinal
lymphomas and its relationship to histological grade S+G2+M phase
fraction ( flow cytometric analysis) and prognosis. Histopathology
1991;18:21-27.
150
75. Muskhelıshvılı L, Wıngard
SK,
Latendresse JR. Proliferating Cell
Nuclear Antigen—A Marker for Ovarian Follicle Counts Toxicologic
Pathology.2005; 33:365–368
76. Oktay K, Schenken RS, Nelson JF. Proliferating cell nuclear antigen
Marks the initiation of follicular growth in the rat. Biol Reprod. 1995;53:
295-301.
77. Greenberg ME, Ziff EB. Stimulation of 3T3 cells induces transcription
of the c-fos proto-oncogene. Nature 1984; 311: 433-438 .
78. Fisch TM, Prywes R, Reoder RG. c-fos sequence necessary for basal
expression and induction by epidermal growth factor, 12-Otetradecanoyl phorbol-13-acetate and the calcium ionophore. Mol Cell
Biol 1987; 7: 3490-3502.
79. Roche E, Buteau J, Aniento I, Reig JA, Soria B, Prentki M. Palmitate
and oleate induce the immediate-early response genes c-fos and nur77 in the pancreatic beta-cell line INS-1. Diabetes 1999; 48: 20072014.
80. Scholf C, Waring M, Bergwitz C, Arseniev L, von zur Muhlen A,
Brabant G. Cyclic-adenosine 3‘,5‘-monophosphate-stimulated c-fos
gene transcription involves distinct calcium pathways in single betacells. Mol Cell Endocrinol 2002; 186: 121-131.
81. Zhu YS, Brodsky M, Franklin SO, Huang T, Inturrisi CE. Metrazole
induces the sequential activation of c-fos, proenkephalin, and tyrosine
hydroxylase gene expression in the rat adrenal gland: modulation by
glucocorticoid and adrenocorticotropic hormone. Mol Pharmacol 1993;
44: 328-335.
82. Susini S, Roche E, Prentki M, Schlegel W. Glucose and glucoincretin
peptides synergize to induce c-fos, c-jun, junB, zif-268, and nur-77
gene expression in pancreatic ß (INS-1) cells. The FASEB Journal.
1998; 12: 1173-1182.
151
83. Stachowiak MK, Sar M, Tuominen RK, Jiang HK, An S, Iadarola MJ,
Poisner A, Hong JS. Stimulation of adrenal medullary cells in vivo and
in vitro induces expression of c-fos proto-oncogene. Oncogene 1990;
5: 69-73.
84. Iwata K, Takahashi O, Tsuboi Y, Ochiai H, Hibiya J, Sakaki T,
Yamaguchi Y, Sumino R. Fos protein induction in the medullary dorsal
horn and first segment of the spinal cord by tooth-pulp stimulation in
cats. Pain 1998; 75: 27-36
85. Glover JNM, Harrison SC. Crystal structure of the Heterodimeric bZIP
Transcription Factor c-Fos/ c-Jun Bound to DNA. Nature 1995; 373:
257-261.
86. Nankova BB, Rivkin M, Kelz M, Nestler EJ, Sabban EL. Fos-related
antigen 2: potential mediator of the transcriptional activation in rat
adrenal medulla evoked by repeated immobilization stress. J Neurosci
2000; 20: 5647-5653
87. Fleischmann A, Hafezi F, Elliott C, Reme EC, Rüther U, Wagner FE.
Fra-1 replaces c-Fos-dependent functions in mice. Gene and Dev
2000; 14: 2695-2700
88. Fralix KD, Zhao S, Venkatasubbarao K, Freeman JW. Rap1 reverses
transcriptional repression of TGF-ß Type II receptor by a mechanism
involving AP-1 in the human pancreatic cancer cell line, UK Pan-1.
Journal of Cellular Physiology 2002; 194: 88-98
89. Kool J, Hamdi M, Cornelissen-Steijger P, van der Eb AJ, Terleth C,
van Dam H. Induction of ATF3 by ionizing radiation is mediated via a
signaling pathway that includes ATM, Nibrin1, stress-induced MAP
kinases and ATF-2. Oncogene. 2003; 22: 4235-4242.
90. Stamp JA, Herber J. Multiple immediate-early gene expression during
physiological
and
endocrine
adaptation
to
repeated
stress.
Neuroscience 1999; 94: 1313-1322
152
91. Franchi A, Calzolari A, Zampi G. Immunohistochemical detection of cfos and c-jun expression and cartilaginous tumours of the skeleton.
Virchows Arch 1998; 432: 515-519.
92. Angel P, Karin M . The role of Jun, Fos and the AP-1 complex in cellproliferation and transformation. Biochem Biophys Acta 1991; 1072:
129-157
93. Karin M, Zhend-gang L, Zandi E. AP-1 function and regulation. Curr
Opin Cell Biol 1997; 9: 240-246
94. Foletta VC. Transcription factor AP-1, and the role of Fra-2. Immunol
Cell Biol 1996; 74: 121-133
95. Bakiri L, Matsuo K, Wisniewvska M, Wagner EF, Yaniv M. Promoter
specificity and biological activity of tethered AP-1 dimers. Mol Cell Biol
2002; 22: 4952-4964.
96. Chiarini LB, Freitas FG, Petrs-Silva H, Linden R. Evidence that the
bifunctional redox factor/AP endonuclease Ref-1 is an anti-apoptotic
protein associated with differentiation in the developing retina. Cell
Death Differ 2000; 7: 272-281
97.Nishikawa S. Localization of transcription factor AP-1 family proteins in
ameloblast nuclei of the rat incisor. Journal of Histochemistry and
Cytochemistry 2000; 48: 1511-1520
98.
De
Sousa
SO,
Mesquita
RA,
Pinto
DS
Jr,
Gutkind
S.
Immunolocalization of c-Fos and c-Jun in human oral mucosa and in
oral squamous cell carcinoma. J Oral Pathol Med 2002; 31: 78-81
99. Lee CS, Charalambous D. Immunohistochemical localisation of the cfos oncoprotein in pancreatic cancer. Zentralbl Pathol 1994; 140: 271275
100. Roivainen A, Söderström KO, Pirila L, Aro H, Kortekangas P, Palo
MR, Jama YT, Toivanen A, Toivanen P. Oncoprotein expression in
human synovial tissue: an immunohistochemical study of different
types of arthritis. British Journal of Rheumatology 1996; 35: 933-942
153
101. Ahn S, Olive M, Aggarwal S, Krylov D, Ginty DD, Vinson C. A
dominant-negative inhibitor of CREB reveals that is general mediator
of stimulus-dependent transcription of c-fos. Mol Cell Biol 1998; 18:
967-977.
102. Oshitari T, Dezava M, Okada S, Takano M, Negishi H, Horie H,
Sawada H, Tokuhisa T, Adachi-Usami E. The role of c-fos in cell
death and regeneration of retinal ganglion cells. Invest Ophthalmol
Vis Sci 2002; 43: 2442-2449.
103. Morgan JI, Curran T. Immediate-early genes: ten years on. Trends
Neurosci 1995; 18: 66-67.
104. Skar R, Larsen TH, Serck-Hanssen G. Regulation of c-fos expression
by IGF-I in bovine chromaffin cells: desensitization following
cholinergic activation. Mol Cell Endocrinol 1994; 106: 213-220.
105. Shea-Eaton W, Sandhoff TW, Lopez D, Hales DB, McLean MP.
Transcriptional repression of the rat steroidogenic acute regulatory
(StAR) protein gene by the AP-1 family member c-Fos. Mol Cell
Endocrinol 2002;188:161–170.
106. Jo Y, King SR, Khan SA, StoccoDM. Involvement of protein kinase C
and cyclic adenosine 3_,5_-monophosphate-dependent kinase in
steroidogenic acute regulatory protein expression and steroid
biosynthesis in Leydig cells. Biol Reprod 2005;73:244–255.
107. Victor E. Beshay, Jon C. Havelock, Rosa Sirianni, Ping Ye, Takashi
Suzuki, William E. Rainey, Bruce R. Carr. The Journal of Clinical
Endocrinology & Metabolism 2007; 92(12):4802–4809.
108. Williams RL, Hilton DJ, Pease S. Myeloid leukaemia inhibitory factor
maintains the development potential o embriyonic stem cells. Nature
1988; 336: 684-687.
109. Matsui, Y., Toksoz, D., Nishikawa, S., et al. (1991) Leukaemia
inhibitor factor on mutine primordial germ cells in culture.Nature, 353,
750–752.
154
110. Yamamori, T., Fukada, K., Aebersold, R. et al. (1989) The cholinergic
neuronal differentiation factor from heart cells is identical to leukemia
inhibitoryfactor (published erratum appears in Science, Jan 19, 1990,
271, 4940).Science, 246(4936), 1412–1416.
111. Murphy, M., Reid, K., Hilton, D.J. and Bartlett, P.F. (1991) Generation
of sensory neurons is stimulated by leukemia inhibitory factor. Proc.
Natl. Acad. Sci. USA, 88(8), 3498–3501.
112. Reid, L.R., Lowe, C., Cornish, J. et al. (1990) Leukemia inhibitory
factor: a novel bone-active cytokine. Endocrinology, 126(3), 1416–
1420.
113. Mori, M., Yamaguchi, K. and Abe, K. (1989) Purification of a
lipoprotein lipase-inhibiting protein produced by a melanoma cell line
associated with cancer cachexia. Biochem. Biophys. Res. Comm.,
160(3), 1085–1092.
114. Baumann, H. and Wong, G.G. (1989) Hepatocyte-stimulating factor
III shares structural and functional identity with leukemia-inhibitory
factor. J. Immunol., 143(4), 1163–1167
115. Smith ER, Xu X. Etiology of epithelial ovarian cancer: a cellular
mechanism for the role of gonadotropins. Gynecol Oncol 2003;91:1–
2.
116. Cramer DW, Welch WR. Determinants of ovarian cancer risk. II.
Inferences regarding pathogenesis. J Natl Cancer Inst 1983;71:717–
721.
117. Nieto JJ, Crow J, Sundaresan M et al. Ovarian epithelial dysplasia in
relation to ovulation induction and nulliparity. Gynecol Oncol
2001;82:344–349.
118. Rossing MA, Daling JR, Weiss NS, Moore DE, Self SG. Ovarian
tumors in a cohort of infertile women. N Engl J Med 1994;331:771–
776.
155
119. Shushan A, Paltiel O, Iscovich J, Elchalal U, Peretz T, Schenker JG.
Human menopausal gonadotropin and the risk of epithelial ovarian
cancer. Fertil Steril 1996;65:13–8.
120. Harris R, Whittemore AS, Itynre J. The collaborative Ovarian Cancer
Group. Characteristics relating to ovarian cancer risk: collaborative
analysis of 12 US case control studies. III. Epithelial tumors of low
malignant potential in white women. Am J Epidemiol 1992;136:
1204–1211.
121. Chene G, Penault –Llorca F, Le Bouedec G, Mishellany F, Dauplat
MM, Jaffeux P, Aublet-Cuvelier B, Pouly JL, Dechelotte P, Dauplat J.
Ovarian epithelial dysplasia after ovulation induction: time and dose
effect. Human Repro 2009;24:132-138.
122. Celik C, Gezginc K, Aktan M, Acar M, Yaman ST, Gungor S, Akyurek
C. Effects of ovulation induction on ovarian morphology: an animal
study. Int Gynecol Cancer 2004;14:600-606.
123. Sushan A, Paltiel O, Schencker JG. Induction of ovulation and
borderline ovarian cancer- the hormonal connection?. Eur J Obstet
Gynecol Reprod Biol 1999;85:1-4.
124. Parazzini F, Negri E, La Vecchia C, Moroni S, Polatti A, Chiaffarino F.
Treatment for infertility and risk of ovarian tumors of borderline
malignancy. Gynecol Oncol 1998; 68: 226-228.
125. Ness RB, Cramer DW, Goodman MT,. Infertility, fertility drugs and
ovarian cancer: a pooled analysis of case- control studies. Am J
Epidemial 2002;155:217-224.
126. Mahdavi A, Pejovic T, Nezhat F. Induction of ovulation induction and
ovarian cancer: a critical review of the literature, fertil Steril
2006;85:819-826.
127. Olivia L. Tan, Peter. R. Hurst, Jean. S. Fleming. Location of inclusion
cysts in mouse ovaries in relation to age, pregnancy, and total
ovulation number: implications for ovarian cancer?. The Journal of
Pathology 2005;205:483–490
156
128. Lin-Zhi Zhuang, Eli Y. Adashi and Aaron J.W. Hsueh . Direct
enhancement
of
gonadotropin-stimulated
ovarıan
estrogen
biosynthesis by estrogen and clomiphene citrate. 1982; 110: 22192221.
129. Williams SC, Gibbons WE, Muasher SJ, Oehninger S. Minimal
ovarian hyperstimulation for in vitro fertilization using sequential
clomiphene citrate and gonadotropin with or without the addition of a
gonadotropin-releasing hormone antagonist. Fertil Steril. 2002
;78:1068-1072.
130. Ernest Hung Yu Ng1, Oi Shan Tang,Pak Chung Ho. The significance
of the number of antral follicles prior to stimulation in predicting
ovarian responses in an IVF programme. Hum. Reprod. 2000; 15 :
1937-1942.
131. A Lass, J Skull, E McVeigh, R Margara ,R M Winston. Measurement
of ovarian volume by transvaginal sonography before ovulation
induction with human menopausal gonadotrophin
for in-vitro
fertilization can predict poor response. Hum. Reprod. (1997) 12 (2):
294-297.
132. Filicori M, Cognigni GE, Taraborrelli S, Spettoli D, Ciampaglia W, de
Fatis CT, Pocognoli P. Luteinizing hormone activity supplementation
enhances
follicle-stimulating
hormone
efficacy
and
improves
ovulation induction outcome. J Clin Endocrinol Metab. 1999 Aug; 84 (
8 ) : 2659 -2663.
133. Willis DS, Watson H, Mason HD, Galea R, Brincat M, Franks S.
Premature response to luteinizing hormone of granulosa cells from
anovulatory women with polycyctic ovary syndrome: revelance to
mechanism of anovulation. J Clin Endocrinol Metab. 1999, 83: 39843991).
157
134. Filicori M, Cognigni G, Tabarelli C, Pocognoli P, Taraborelli S,
Spettoli D, Ciampaglia W. Stimulation and growth of antral ovarian
follicles by selective LH activity administration in women. J Clin
Endocrinol Metab. 2002 ;87(3):1156 -61.)
135. Imani B, Eijkemans MJ, te Velde ER, Habberna JD, Fauser BC.
Predictiors of chances to conceive in ovulatory patients during
clomiphene citrate induction of ovulation in normogonadotropic
oligoamenorrheic infertility. J Clin Endocrinol. Metab 1999;84:16171622.
136. Fatemi HM, Kolibianakis E, Tournaye H, Camus M, Van Steirteghem
AC, Devroey P. Clomiphene citrate versus letrozole for ovarian
stimulation: a pilot study. Centre for Reproductive Medicine, DutchSpeaking Free University Brussels, Laarbeeklaan 101, 1090
Brussels, Belgium 2003; 17:543-546
137. Imani B, Eijkemans MJ, te Velde ER, Habberna JD, Fauser BC.
Predictiors of chances to conceive in ovulatory patients during
clomiphene citrate induction of ovulation in normogonadotropic
oligoamenorrheic infertility. J Clin Endocrinol. Metab 1999;84:16171622.
138. N. Bayram, MV. Wely, FV. Veen. Recombinant FSH versus urinary
gonadotrophins or recombinant FSH for ovulation induction in
subfertility associated with polycystic ovary syndrome. 2001
139. Feranil JB, Isobe N, Nakao T. Cell proliferation in the atretic follicles
of buffalo and cattle ovary. Reprod Domest Anim. 2004; 39: 405-409.
140. Picut CA, Swanson CL, Scully KL, Roseman VC, Parker RF, Remick
AK. Ovarian follicle counts using proliferating cell nuclear antigen
(PCNA) and semi-automated image analysis in rats. Toxicol Pathol.
2008;36:674-679.
158
141. S Mahner, C Baasch, J Schwarz, S Hein, L Wölber, F Jänicke and
K Milde-Langosch-Fos expression is a molecular predictor of
progression and survival in epithelial ovarian carcinoma. British
Journal of Cancer 2008; 99: 1269-1275.
142. Suzanne Roffler-Tarlov1, Jeremy J. Gibson Brown2,4, E. Tarlov3,
Javor Stolarov4, Deborah L. Chapman4,Maria Alexiou4 and Virginia
E. Papaioannou2,4. Programmed cell death in the absence of c-Fos
and c-Jun. Development 1996; 122: , 1-9
143. C. Sharma, JS. Richards. Regulation of AP1 (Jun/Fos) factor
expression and activation in ovarian granulosa cells. Relation of
JunD and Fra2 to terminal differentiation. J Biol Chem. 2000; 275:
33718-28.
144. Kauma, S.W. and Matt, D.W. (1995) Coculture cells that express
leukemia
inhibitory
factor
(LIF)
enhance
mouse
blastocyst
development in vitro. J. Assist. Reprod. Genet., 12, 153–156.
145. Lavranos, T. and Seamark, R. (1989) Myeloid leukemia inhibitory
factor (LIF): an embryotrophic factor. Proc. Aust. Soc. Reprod. Biol.,
21, 91.
146. Fry, R. (1992) The effect of leukemia inhibitory factor (LIF) on
embryogenesis. Reprod. Fertil. Dev., 4, 449–458.)
147. R. Abir, B. Fisch,S. Jin, M. Barnnet, S. Freimann, R. Van den Hurk,
D. Feldberg, S. Nitke ,H. Krissi ,A. A. Immunocytochemical detection
and RT–PCR expression of leukaemia inhibitory factor and its
receptor in human fetal and adult ovaries Mol. Hum. Reprod. (2004) 10
(5): 313-319.
148. A. Arıcı, E. Oral, O. Bahtiyar, O. Engin, E. Seli, EE. Jones.
Leukaemia inhibitory factor Express,on in human follicular fluid and
ovarian cells. Hum Repr 1997;12:1233-1239.
159
10.EKLER
160
11. TEŞEKKÜR
Yüksek lisans eğitimim boyunca bilgi ve deneyimleriyle bana
yol gösteren ve her zaman desteğini gördüğüm tez danıĢmanım ve
Anabilim Dalı BaĢkanımız Prof. Dr. Deniz ERDOĞAN baĢta olmak üzere
katkılarından dolayı Prof. Dr. M. Tahir HATĠPOĞLU‘na, Prof. Dr. Celal
ILGAZ‘a, Prof.Dr. Candan ÖZOĞUL‘a, Prof. Dr. Suna ÖMEROĞLU‘na,
Doç. Dr. Gülnur TAKE KAPLANOĞLU‘na ve Doç. Dr. Çiğdem ELMAS‘a
sonsuz teĢekkürlerimi sunarım.
Tezimin
bütün
aĢamalarında
benden
yardımlarını
esirgemeyen ve her zaman yanımda olup, bana destek olan ArĢ. Gör.
Güleser GÖKTAġ‘a ve Uzman Dr. Fatma HELVACIOĞLU‘na, bütün
dönem arkadaĢlarıma ve tüm hayatım boyunca maddi ve manevi
desteklerini esirgemeyen sevgili annem, babam ve kardeĢime sonsuz
teĢekkür ediyorum.
Sinem DEMİR
Aralık 2010
161
12. ÖZGEÇMİŞ
1985 yılında Ankara‘da doğdum. Ġlköğrenimimi YükseliĢ
Koleji‘nde, orta öğrenim ve liseyi Ankara Üniversitesi GeliĢtirme Vakfı Özel
Lisesi‘nde tamamladım. 2003 yılında Gazi Üniversitesi Fen-Edebiyat
Fakültesi Biyoloji Bölümü‘ne girdim ve 2007 yılında mezun oldum. 2008
yılında Gazi Üniversitesi Tıp Fakültesi‘nde Sağlık Bilimleri Enstitüsü‘nün
Histoloji ve Embriyoloji Anabilim Dalı‘nda yüksek lisans eğitimine
baĢladım.
162