HÜCRE ZARLARINDA
ETKİLEŞMELER
ve ENZİMATİK
TEPKİMELER
Prof. Dr. Hamdi ÖĞÜŞ
[email protected]
Yakın Doğu Üniversitesi Tıp Fakültesi
Biyokimya Anabilim Dalı, Lefkoşa, KKTC
Basit diffüzyon
Eşit olmayan derişimlerde çözünür madde ya da iyon içeren iki
bölme geçirgen bir zarla ayrılmış olsun :
Çözüt (solute), derişimin yüksek olduğu bölmeden düşük
derişimli bölmeye basit diffüzyonla hareket eder.
Bu göç her iki bölmedeki derişim eşitleninceye kadar sürer.
Yüksüz moleküllerin hareketi,
dengeye ulaşılıncaya kadar
düşük derişimli olan tarafa
doğrudur.
Elektriksel yükü olan çözütler ise
hem elektriksel potansiyel, hem de
kimyasal derişimin bir bir bileşimine
uygun olarak hareket ederler
• Biyolojik olarak önemli bazı gazlar basit diffüzyonla zarları
geçebilir : O2, C02, azot, metan.
• Su bazı biyolojik zarları basit diffüzyonla geçebilir.
ZAR TRANSPORT SİSTEMLERİ
Basit diffüzyon
(yalnız nonpolar bileşikler,
elektrokimyasal
gradiyent
yönünde)
İyonofor aracılı
iyon transportu
(elektrokimyasal
gradiyent
yönünde)
İyon kanalı
(elektrokimyasal
Gradiyent yönünde,
iyon ya da ligand ile açılır)
Kolaylaştırılmış diffüzyon
(elektrokimyasal
gradiyent
yönünde)
Primer aktif
transport
(elektrokimyasal
gradiyente karşı)
Sekonder aktif transport
(elektrokimyasal gradiyente
karşı, iyon gradiyenti ile
yürütülür)
Pasif transport (Kolaylaştırılmış diffüzyon)
• Zarlardan geçiş için başka bir
seçenek de pasif transporttur
(kolaylaştırılmış diffüzyon).
• Taşıyıcılar (permeazlar) bir
çözütün zarı geçmesini
kolaylaştıran zar proteinleridir.
• Taşıyıcılar polar bileşiklerin
ve iyonların taşınması için
gerekli aktivasyon enerjisini
düşürürler.
• Transport proteinleri
I) Taşıyıcı proteinler
II) Kanal proteinleri
- Yüksek hız
- Yüksek özgüllük
- Enerji gerektirmez
Pasif transport (Kolaylaştırılmış diffüzyon)
• Polar ya da yüklü bir molekülün ikili
Serbest enerji, G
Islanmış
çözüt
Taşıyıcısız basit
diffüzyon
•
Kolaylaştırılmış
diffüzyon
basit
diffüzyon
•
taşınım
•
•
Taşıyıcı
bir lipid zardan geçebilmesi için,
önce ıslanma kabuğundaki
(hydration shell) su moleküllerinden kurtulması ve sonra zardan
diffüze olması gerekir.
Saf lipid zarlar, polar ve yüklü
moleküllere gerçekten
geçirimsizdir.
Basit diffüzyonda, polar bir çözütün
ikili zardan geçebilmesi için yüksek
bir enerji (Gǂ) gerekmektedir.
Taşıyıcı bir protein, hidrofilik bir zaraşırı geçit sağlayarak gerekli enerjiyi
düşürür.
Bunu, tepkime sırasında enzimlerin
aktivasyon enerjisini düşürmelerine
benzetebiliriz.
Taşıyıcı-aracılı diffüzyon (kolaylaştırılmış diffüzyon) ile pasif
diffüzyonun karşılaştırılması
• Pasif diffüzyonda hareket hızı
çözüt derişimi ile doğru
orantılıdır.
• Taşıyıcı-aracılı diffüzyonda ise
hız doygunluğa erişir.
• Maksimum hızın yarısına
(Vm/2) ulaşmak için gerekli
çözüt derişimi, taşıyıcının bu
molekül için afinitesini ifade
eden bağlanma sabitidir (Km).
• Kolaylaştırılmış diffüzyonun
kinetiği enzimle katalize
edilen tepkime kinetiği ile
benzerdir.
Taşıma sistemlerinde genel sınıflandırma
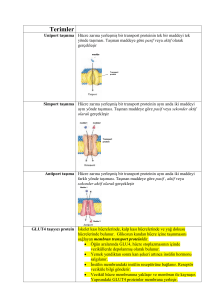
Uniport
Simport
Antiport
Kotransport
Uniport
: Yalnızca bir substrat taşınır (örn. glukozun eritrosit glukoz
taşıyıcısı ile taşınması).
Simport : İki substrat birlikte ve aynı yönde hareket ederler.
Antiport : İki substrat farklı yönlerde hareket ederler.
•
•
•
•
•
•
•
•
•
Kanal proteinleri:
Zar-aşırı taşınım hareketlerinin
taşıyıcı proteinlerinde
olduğundan daha yüksek hızda
olmasına izin verirler.
Stereospesifik özellikleri
taşıyıcı proteinlerinden daha
azdır.
Genellikle taşıma hızı
doygunluğa erişmez.
Oligomerik kompleksler
halindedirler.
Taşıyıcı proteinler (carriers) :
Substratlarına çoklu kovalan etkileşmelerle yüksek düzeyde
stereospesifik olarak bağlanırlar.
Taşınımı serbest diffüzyon sınırlarının altında hızlarla katalize ederler.
Hızları doygunluğa erişir.
Monomerik proteinlerdir.
-heliks yapısında bölgeler ya da -fıçı yapısı içeren proteinlerdir.
Eritrosit zarında klorür ve bikarbonat iyonları, bir
antiport sistemiyle, birlikte fakat ters yönlerde taşınırlar
• Eritrosit dokulardan akciğerlere CO2
Bikarbonat kan
plazmasından
eritrosite girer.
taşınması için temel olan bir anyon
değiştirici, kolaylaştırılmış diffüzyon sistemi
içerir.
• Atık CO2 dokulardan eritrositlere salınır,
karbonik anhidraz ile hemen bikarbonata
(HCO3) çevrilir (HCO3- kan pH’sının birincil
tamponudur).
• Bikarbonat kan plazmasına yeniden girer ve
akciğerlere taşınır.
• Bikarbonat kan plazmasında CO2 ’den çok
daha fazla çözündüğünden kanın CO2
taşıma kapasitesi artar.
• Akciğerlerde bikarbonat tekrar aynı enzimle
CO2‘ye çevrilir ve nefesle atılır.
• Eritrositler AE1 tipi taşıyıcı içerirken, AE2
tipi K.C.’de, AE3 ise beyin, kalp ve retinada
baskın tiptir.
GLUKOZ TAŞIYICISI GLUT-1 İYİ BİR ÜNİPORT ÖRNEĞİDİR.
• Glukoz eritrosite, özgül bir glukoz taşıyıcı yardımıyla (kolaylaştırılmış
diffüzyonla) ve katalize edilmemiş pasif diffüzyondan yaklaşık
5000 kez daha hızlı olarak girer.
• Eritrosit glukoz taşıyıcısı (GLUT1) bir tip III integral proteindir.
Molekül ağırlığı 45000 olan bu taşıyıcı, zarı geçen ve heliks yapan
yapıda 12 hidrofobik segmente sahiptir.
Hidrofobik
Polar
Yüklü
Dış
İç
GLUKOZ TAŞIYICISI GLUT-1 İYİ BİR ÜNİPORT ÖRNEĞİDİR.
Eritrositte glukoz transportu
Taşıyıcı iki konformasyonda bulunur:
T1
Taşıyıcının glukoz bağlayan bölgesinin
plazma zarının dış yüzeyinde olduğu
konformasyon
T2
Bağlanma bölgesinin iç yüzeyde
olduğu konformasyon
Glukoz taşıyıcı dört aşamada görülür:
1)Kan plazmasındaki glukoz T1 konformasyonunda stereospesifik olan bir bölgeye
bağlanır. Bu aktivasyon enerjisini düşürür.
2)Konformasyon T1’den T2’ye değişir.
3)Glukoz T2’den sitoplazmaya salıverilir.
4)
Taşıyıcı T1 konformasyonuna döner.
Artık yeni bir glukoz molekülünü taşımaya
hazırdır.
GLUKOZ TAŞIYICILAR (GLUT ailesi)
Taşıyıcı
GLUT1
GLUT2
GLUT3
GLUT4
Diğerleri
Dağılımı
Açıklama
Eritrosit, endotel, kanbeyin barajı, glia ve diğer
hücreler
Böbrek tübüler epitel h.,
İ.B. epitel h., K.C. ve
pankreatik β- hücreleri
Nöronlar ve plasenta
Kan glukoz düzeyine göre
düzenlenirler
Yağ dokusu ve çizgili kas
dokusu (kalp ve iskelet
kası)
İnsülin-bağımlı glukoz taşıyıcı.
Glukozun insülinle düzen-lenen
depolanmasından sorumlu
Yüksek kapasite, düşük afinite
Çok yüksek kapasiteli izoform
GLUT 5 (fruktoz taşıyıcı), GLUT 6, GLUT 7 (endoplazmik
retikülumdan dışarı doğru), GLUT 8-12.
AKTİF TRANSPORT
• Transportun yönü, çözütün derişimine ya da
elektrokimyasal gradiyente karşıdır.
• Bir taşınım proteini gerektirir (daima taşıyıcı
proteinler, kanal proteinleri değil)
• Enerji girdisi gereklidir
• - ATP hidrolizi (primer aktif transport)
- Elektrokimyasal gradiyent yönünde akarak eşlik
eden bazı iyonlarla (sekonder aktif transport)
İKİ TÜR AKTİF TRANSPORT VARDIR
• Primer aktif transportta,
ATP hidrolizinden ortaya
çıkan enerji çözütün
elektrokimyasal gradiyente
karşı taşınması için ve
doğrudan kullanılır.
• Sekonder aktif transportta
ise, primer aktif transportla
yaratılan bir X iyonu
gradiyenti vardır.
• X’in elektrokimyasal gradiyenti yönünde (yüksek derişimden
düşük olan tarafa) hareketi, ikinci çözütün (S) kendi elektrokimyasal gradiyentine karşı göçünü sağlayan sürücü güçtür.
SEKONDER AKTİF TRANSPORT EMİLİMDE ÖNEMLİDİR
İnce bağırsak ve böbrekte glukoz transportu
Na+-glukoz taşıyıcılar (simport)
Glukoz, Na+ ile birlikte apikal
yüzdeki zardan epitel hücresine
taşınır (simport).
Hücrenin bazal yüzeyine gelen
glukoz burada pasif bir taşıyıcı
olan GLUT2 yardımıyla kana
geçer (uniport).
Na+ ise Na+,K+-ATPaz tarafından
yine bazal yüzden hücre dışına,
K+ ise aynı yüzden hücre içine
pompalanır.
Bu sistemin yürütücü gücü Na+ gradiyentidir. Bu gradiyent Na+/K+ATPaz sistemi tarafından, ATP hidrolizinden sağlanan enerji kullanılarak oluşturulduğundan, ince bağırsaktaki (ve böbrek tüp
epitelindeki) glukoz transportu sekonder aktif transport tipindedir.
ATP kullanan Aktif Transport Sistemleri
• P-sınıfı pompalar → Bir altbirimde aspartat yan grubunun
fosforilasyonu söz konusudur.
• F- ve V-sınıfı pompalar → Fosfoprotein yapısında bir
arabileşik yoktur, yalnızca proton transportu var.
• ABC (ATP-binding cassette) üstailesi.
ATP kullanan Aktif Transport Sistemleri
• P-tipi ATPazlar :
Na+/K+ ATPaz
H+/K+ ATPaz
Ca+ ATPaz
İyonlar zıt yönlerde hareket ederler.
: Plazma zarında bulunurlar
: Plazma zarında bulunurlar
: Plazma zarında ve endoplazmik
retikülumda bulunurlar.
• V-tipi ATPazlar: İyonlar bir yönde hareket ederler.
H+ : Lizozomal ve endozomal keseciklerde bulunurlar
• F-tipi ATPazlar: İyonlar bir yönde hareket ederler.
H+ : Mitokondri iç zarında bulunurlar.
• Çoklu-ilaç taşıyıcılar (Multidrug transporter)
Tümör hücrelerinin plazma zarlarında bulunurlar.
ATP kullanan Aktif Transport Sistemleri
•
•
•
•
P-tipi ATPazlar
ATP-bağımlı aktif katyon taşıyıcılar tersinir olarak fosforile
olurlar.
Fosforilasyon proteini bir konformasyon değişikliğine
zorlayarak katyonun zardan geçebileceği bir ortam yaratır.
Bütün P-tipi transport ATP’ları benzer amino asit dizilerine
sahiptirler. Proteindeki bir Asp yan grubu forforile olur.
Bütün P-tipi ATPaz’lar bir fosfat analoğu olan vanadat ile
inhibe olurlar.
Na+-K+-ATPaz (Na-K pompası)
• Na+,K+ -ATPaz, bütün hücrelerin
ana iyon pompasıdır. Bu pompa
olmadan yaşam olanaksızdır.
• Hücre içinde ve dışında yer alan
iyonların derişimini hem kimyasal
hem de elektrokimyasal gradiyent
olarak dengelenme ve diğer
transport sistemlerine de yardımcı
yardımcı olma görevi vardır.
• Na+,K+ -ATPaz, her ikisi de zarı aşan
iki altbirime (α ve ) sahip bir
integral proteindir.
• Na+ ve K+ için bir antiport sistemidir.
• ADP ve Pi’ye çevrilen her ATP molekülü için, iyonlar plazma zarından
içeri (2 K+) ve dışarı (3 Na+)
pompalanır.
Na+-K+-ATPaz POMPASININ ÇALIŞMA MEKANİZMASI
Na-K ATPaz iki form arasında döngü yapar
• Fosforile formu (P-Enz-II) K+’a
•
•
•
•
yüksek, Na+’a ise düşük afinite
gösterir
Defosforile formu (P-Enz-II) ise
Na+’a yüksek, K+’a ise düşük
afinite gösterir
İçeri giren her 2 K+ iyonuna karşılık
3 Na+ iyonu dışarı çıkar.
Süreç zarın iki tarafında net bir yük
ayırımı yapar.
Sonuçta -50 ile -70 mV arasında
(içeride dışarıya göre negatif) bir
trans-membran potansiyel oluşur
ve bu potansiyel nöronlarda aksiyon
potansiyelinin iletilmesi için esastır.
Na+-K+-ATPaz POMPASININ İNHİBİTÖRLERİ
•
•
•
•
•
Digitoksijenin;
Digitalis glikozidlerinin
aktif bileşenidir,
Konjestif kalp
yetmezliğinde
kullanılır,
Kalp atım hızını
Digitoxigenin
artırmadan kalp
kasının kasılmasını güçlendirir,
Kalbin çalışma verimini artırır.
Etki mekanizması:
Digitalis glikozidleri Na+ çıkışını inhibe eder. Hücre içinde
artan Na+ derişimini kalp kası hücresindeki Na+-Ca2+
antiport sistemini aktive ederek hücreiçi Ca++ derişimini
artırır ve sonuçta kalp kasının kasılmasını güçlendirir.
H+-K+ ATPaz (Proton pompası)
P-tipi bir ATPaz’dır. Memelilerde gastrik mukoza ile sınırlı olan H+K+ ATPaz, mide parietal hücrelerinde karbonik anhidrazla üretilen
hidrojen iyonunu (H+) ATP hidrolizi karşılığında K+ iyonu ile değiştokuş yapar. Klorür iyonu da potasyum ile birlikte atılır.
Sarkoplazmik Retikülum Kalsiyum Pompası (Ca++-ATPaz)
• Serbest kalsiyumun (Ca2+) sitozoldeki
derişimi genellikle 100 nM’dan daha
düşüktür.
• Kalsiyum iyonları sitozol dışına plazma
zarı kalsiyum pompası ile pompalanır.
• ER zarındaki Ca2 pompası kalsiyumu
sitoplazmadan ER lümenine pompalar.
• Kas hücrelerinde (miyosit), Ca2+
özelleşmiş bir ER’de (sarkoplazmik
retikülum) hapsedilir.
• Sarkoplazmik ve endoplazmik
retikülum kalsiyum pompaları
SERCA pompaları olarak anılırlar.
- Plazma zarı Ca2 pompası Fosforile/defosforile konformasyonlar
arasında döngü yapan integral zar
- SERCA pompaları
proteinleridir.
V-tipi ATPaz (Vakuoler ATPaz, V-ATPaz)
• V-ATPaz, mitokondri iç zarında yer
ATP
H+ ADP + Pi
V-ATPaz
Sitoplazma
H+
* Kesecik
H+
H+
Ca2+
Ca2+
alan FoF1-ATPaz yapısına benzer bir
yapıdadır.
• Görevi, yerleştiği hücreiçi organellere ATP hidroliz enerjisini
kullanarak sitoplazmadan H+
pompalamaktır.
• Oluşan hidrojen iyonu gradiyenti
daha sonra sekonder aktif transportla (genellikle antiport)
organele iyon ya da molekül
alımımda kullanılır.
* Kesecik; lizozom, endozom, diğer
hücre organelleri olabilir. Bu örnekte,
sitoplazmik kalsiyum keseciklerde
toplanmaktadır.
F - tipi ATPaz (ATP sentaz)
Mitokondride, bakteride ve kloroplastta tepkimelerden sağlanan
enerjiden ATP sentezi için kullanan bir aktif transport sistemidir.
F1 : periferal bölge: 3
altbirimi, 3 altbirimi, 1
altbirimi (mor), ve merkezi bir
eksen (, yeşil altbirim).
F0 : integral bölge; c, 1 a, ve 2
b altbirimlerinin birçok
kopyası bulunur (İnhibitör:
oligomisin).
ATP enerjisini kullanan F1 , bir moleküler bir motor gibi çalışır,
hidrolize ettiği her ATP molekülü için (4) protonu, daha
derişik bulundukları bir ortama pompalar.
Fo , F1 tarafından pompalanan protonlar için zaraşırı
(transmembrane) bir kanal oluşturur.
ATP’yi sentezleyen ve yıkan (ATPaz) aktiviteler
F1 bölgesinde yer alır
F-tipi ATPazlarla katalize
edilen tepkimeler
tersinirdir, bir proton
gradiyenti tepkime
yönünü tersine çevirecek
(ATP sentezi ) enerjiyi
sağlayabilir.
Bu biçimde ters yönde
çalışan ATPazlar (ETZ,
kompleks V) daha uygun
bir deyimle, ATP sentaz
olarak anılırlar.
Çoklu-ilaç taşıyıcılar (ABC transporters)
• MDR1, zaraşan 12 bölgesi
ve iki ATP bağlayıcı bölgesi
(kaset) olan bir integral zar
proteinidir (Mr 170,000).
• Bu taşıyıcı protein ailesi,
ABC taşıyıcı ailesi
(ATP-Binding Cassette,
ABC transporter) olarak
anılmaktadır.
• ABC taşıyıcılar, amino asitler, peptidler, proteinler, metal
iyonları, çeşitli lipidler ve ilaçları da içeren birçok hidrofobik
bileşikleri hücre dışına ve yüksek derişime karşı taşıyan
ATP-bağımlı taşıma proteinleridir (aktif transport).
Çoklu-ilaç taşıyıcılar (ABC transporters)
Çoklu-ilaç direnç proteini (MDR1,
multidrug resistance protein 1)
insanlarda da bulunan ve tümörlerin bazı etkili antikanser ilaçlarına
direnç göstermesinden sorumlu bir
ABC taşıyıcı sistemidir.
“P-glikoprotein” olarak da bilinir.
Dış
İç
Bu taşıyıcılar, ilaçları
tümör hücresi dışına
pompalayarak hücrede
birikmesini önlerler ve
tedaviyi engellerler.
İyonlar, proteinler tarafından oluşturulan iyon
kanalları aracılığıyla zarlardan geçebilirler
1. Kanallardan akış hızı taşıyıcılardan daha hızlı olabilir.
2. İyon kanallarından geçiş hızı doygunluğa ulaşmaz.
3. İyon kanalları, bazı hücresel olay ve durumlara göre “açıkkapalı” şeklinde denetlenirler.
a) Voltaj-kapılı iyon kanalları, transmembran elektriksel
potansiyeldeki (Vm) bir değişiklik zarda yerleşen yüklü bir
proteinini hareket ettirerek iyon kanalını açar ya da kapar.
örn. Nöronların ve miyositlerin plazma zarındaki Na+, Ca2+
ve K+ iyon kanalları.
b) Ligand-kapılı kanallar, Hücre dışından ya da içinden küçük
bir molekülün bağlanarak proteinde allosterik bir geçişe
neden olmasıyla açılan ya da kapanan kanallardır.
İyonofor - Zarlardan iyon taşınmasını sağlarlar
•
•
•
•
İyonların yükünü maskeleyen küçük moleküllerdir.
İkili-lipid zardan kolayca diffüzlenmelerini sağlarlar.
Enerji kaynaklarına bağlı değildirler.
Çoklukla mikroorganizmalar tarafından sentezlenirler.
• Peptid yapısında bir iyonofor
olan valinomycin, potasyumu
(K+) bağlar.
• K+-valinomisin kompleksinin dış
kısmı hidrofobiktir. Bu nedenle
zardan kolayca diffüze olur ve
potasyum iyonunu derişim
gradiyenti yönünde karşı tarafa
taşır.
Şekilde; K+ yeşil; oksijen atomları
kırmızı ; hidrofobik amino asitlerin
yan zincirleri sarı olarak
gösterilmiştir.
Nöronların voltaj-kapılı (voltage-gated) Na+ kanalları
• Kanalın α-altbirimi dört homolog
bölge (I-IV) içerir ve her bölgede altı
zar-aşan heliks yapısı vardır (1-6).
• Bu dört bölge bir kanal oluşturacak
şekilde bir araya gelirler . Merkezi
kanal polar amino asitlerle kaplıdır.
• Kanal zar depolarizasyonuna yanıt
olarak açılır.
Asetilkolin Reseptörü ligand-kapılı Bir İyon Kanalıdır
• Voltaj kapılı sodyum kanallarının açılması
o bölgede bir depolarizasyon yaratır.
• Depolarizasyon dalgası akson ucuna
ulaştığında, voltaj kapılı Ca++ kanalları
açılır ve presinaptik nörona kalsiyum
girişine olanak sağlar.
• Sonuçta artan kalsiyum asetilkolin (Ach)
içeren keseciklerin ekzositozla sinaptik
yarığa boşalmasını tetikler.
• Ach post-sinaptik nöronda bulunan
reseptörüne bağlanarak ligand-kapılı
sodyum kanallarının açılmasına neden olur.
• Ekstrasellüler Na+ ve Ca++ bu kanaldan
girerek post-sinaptik hücre zarını
depolarize eder.
• Asetilkolin reseptörü, bir motor nörondan
nöromüsküler plakta bir kas lifine sinyalin
geçebilmesi için esastır.
Birçok “iyon kanalı” inhibitörü ile karşılaşabiliriz.
• Tetrodotoksin (fugu balığı)
voltaj-kapılı Na+ kanallarına
bağlanır ve bloke eder.
Saksitoxin (dinoflagellate →midye)
• Saksitoxin
→midye)
voltaj-kapılı(dinoflagellate
Na+ kanallarına
voltaj-kapılı
Na+ kanallarına
bağlanır ve bloke eder.
bağlanır ve bloke
eder.
• Dendrotoksin (yılan zehiri),
voltage-kapılı K+ kanalları ile etkileşir.
Kistik Fibrozis Transmembran Düzenleyici (CFTR)
• CFTR, aslında bir klorür (Cl-) iyon kanal
proteinidir. Zaraşan 12 heliks yapısı
içerir.
• Sitoplazmik yüzeyde yer alan NBD1 ve
NBD2 ATP-bağlayıcı bölgelerdir.
R(regulatory) bölgesi ise cAMP-bağımlı
protein kinaz tarafından fosforile olan
düzenleyici bölgedir. Dış yüzeyde ise
oligosakkarid zincirleri eklenmiştir.
• En sık olarak NBD1 bölgesinde görülen
mutasyon proteinde bir katlanma
sorununa yol açar. Hastalar özellikle
solunum yollarını tıkayan çok koyu
kıvamlı mukus salgılarlar.
AC enfeksiyonları ve diğer dokulardaki
sorunlar nedeniyle hastalar ancak 30
yaşına kadar yaşarlar.
• CFTR; Cystic Fibrosis Transmembrane
Conductance Regulator
Endositoz
Endositoz
Endositozla
alınan besin
tanecikleri
Çevre
Ekzositozla
atılan atık
Sindirim
ürünleri
sitoplazma
Sindirilmiş
besin
Sekonder
lizozomla
birleşme
• Hücrenin büyük molekülleri içine alabilmek için kullandığı işlem
”endositoz”dur. Bu moleküllerden bazıları (örn. polisakkaridler,
proteinler, polinükleotidler) hidrolize edildiğinde hücrede besin
olarak kullanılır.
• Endositik keseciklerin çoğu asidik hidrolitik enzimleri içeren ve
hücrenin mutfağı olan lizozomlarla birleşir.
ENDOSİTOZ TÜRLERİ
Ekstrasellüler SİTOPLAZMA
sıvı
Besin/diğer
parçacıklar
(a) Fagositoz
Plazma zarı
I) Fagositoz
- Yalnızca makrofaj ve
granülositlerde görülür.
- Virüs, bakteri, hücreler ve
kalıntıları kapsar.
II) Pinositoz
- Bütün hücrelerde görülür.
- Hücrenin sıvı ve içeriklerini
alması sağlar.
(b) Pinositoz
Klatrin
Kaplı
girinti
Kaplı
kesecik
(c) Reseptör-aracılı endositoz
III) Reseptör-aracılı endositoz
Makromoleküllerin klatrinle
kaplı kesecikler yoluyla seçici
olarak alınmasını sağlayan
işlemdir.
EKZOSİTOZ
EKZOSİTOZ
• Ekzositoz için sinyal sıklıkla bir hormon ya da nörotransmitterdir.
• Hücre yüzeyinde reseptörüne bağlandığında, yerel ve geçici bir
Ca2+ derişimi değişikliği yaratır ve bu da ekzositozu tetikler.
LDL METABOLİZMASI - ENDOSİTOZ
Hatalı LDL reseptörü, LDL’nin içeri
alınmasını ve hücre içinde kolesterol
sentezinin inhibisyonunu önler.
Kolesterol
Fosfolipid
Apo B-100
Apo A1
HDL
ABCA1
LDL
res.
Normal
Ailesel hiperkolesterolemi
Kolesterol
Fosfolipid
Apo B-100
Apo A1
Hücre zarında yer alan ABCA1 ( bir
ABC taşıyıcı) mutasyonu nedeniyle
hücreiçi kolesterol HDL’ye aktarılamaz.
Sonuçta “ters kolesterol transportu”
durur.
HDL
ABCA1
LDL
res.
Normal
Tangier (Tanca) Hast.
Akuaporinler (Aquaporin) suyun hücre zarından geçebilmesi
için hidrofilik kanal oluşturan bir integral zar protein ailesidir
Akuaporinlerin yapısı : Homotetramer bir moleküldür. Her
altbirim su molekülleri için bir zar-aşırı bir kanal yapar.
DİABETES İNSİPİDUS : Anormal AQUAPORİN >>> POLİÜRİ
Diabetes insipidus
• Böbrek toplayıcı kanallarını döşeyen epitel hücre
zarlarının su geçirgenliği, bu hücrelerin apikal plazma
zarlarına yerleşmiş bir akuaporin (AQP-2) proteinine
bağlıdır.
• Antidiüretik hormon (ADH, vazopressin) epitel
hücrelerinde sitoplazmada kesecik zarlarında depolanan
AQP-2 moleküllerini harekete geçirilip plazma zarına
aktarılmasını ve suyun emilerek korunmasını sağlar.
• Diabetes insipidus hastalığında, AQP-2 genindeki bir
mutasyon su geri-emiliminde önemli sorunlar yaratır,
hastalar litrelerce çok seyreltik idrar çıkarırlar.
“Diabetes insipidus”un en sık rastlanılan türünde ise;
antidiüretik hormonun sentezi ve hipofizden
salgılanmasında bozukluk vardır.