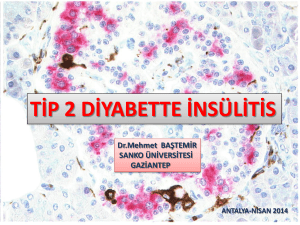
tc selçuk ünġversġtesġ fen bġlġmlerġ enstġtüsü sığırlarda doku
advertisement

T.C. SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ SIĞIRLARDA DOKU SPESĠFĠK GENLERĠN MĠKROARRAY VERĠLERĠNĠN KARġILAġTIRMALI ANALĠZĠYLE BELĠRLENMESĠ Selçuk KAPLAN DOKTORA TEZĠ Zootekni Anabilim Dalı Temmuz-2013 KONYA Her Hakkı Saklıdır ÖZET DOKTORA TEZĠ SIĞIRLARDA DOKU SPESĠFĠK GENLERĠN MĠKROARRAY VERĠLERĠNĠN KARġILAġTIRMALI ANALĠZĠYLE BELĠRLENMESĠ Selçuk KAPLAN Selçuk Üniversitesi Fen Bilimleri Enstitüsü Zootekni Anabilim Dalı DanıĢman: Prof. Dr. Saim BOZTEPE 2013, 62 Sayfa Jüri DanıĢman Prof. Dr. Saim BOZTEPE Prof. Dr. Ramazan YETĠġĠR Prof. Dr. Murat Soner BALCIOGLU Prof. Dr. Birol DAĞ Doç. Dr. Fulya ÖZDĠL Doku spesifik genler geliĢim ve metabolizma da önemli roller oynamaktadır. Günümüzde, gen bankasında farklı dokularla ilgili 3.363.628 GEO Profil ve 397.988 mikroarray verisi bulunmaktadır. Bilim adamları için gen bankasında bu kadar çok faydalı veri bulunmasına rağmen, yeni doku spesifik genlerin ortaya çıkarılması için bu verileri analiz edip kullanmaya yönelik bir strateji izlenmesiyle ilgili eksiklikler bulunmaktadır. Bu çalıĢma, mikroarray ve mikroarraylerle iliĢkili GEO Profillerin kullanımı ve analiziyle doku spesifik genlerin ortaya çıkarılmasına yönelik yeni bir metot ortaya koymaktadır. Bu çalıĢmada, adipoz doku bu metodun doğrulanmasında hedef doku olarak seçilmiĢtir. Ġlk olarak, insan ve fare mikroarray verileri karĢılaĢtırmalı olarak analiz edilmiĢtir. Mikroarray verilerini desteklemek için, adipoz dokuyla ilgili GEO Profilleri gen bankasından seçilmiĢtir. Daha sonra, adipoz dokuyla ilgili microarray ve GEO Profil verileri eĢ zamanlı olarak değerlendirilmiĢtir. Sonuç olarak bu metot, Chordin-like 1 (Chrdl1) olarak isimlendirilen yeni bir adipoz spesifik geni hipotez etmiĢtir. Hipotezin testi için sığır dokularında RT-PCR yapılmıĢtır. RT-PCR sonuçlarına göre, Chrdl1 diğer dokulara göre adipoz dokuda yüksek derecede ekspresyon göstermiĢtir (P<0.05). Bu sonuç, kurulan hipotezi desteklemiĢ ve bu çalıĢmada kullanılan metodun yeni doku spesifik genlerin belirlenmesinde faydalı olabileceğini göstermiĢtir. Anahtar Kelimeler: Adipoz doku, Adipoz spesifik gen, Atgl, Chrdl1, Fabp4, Mikroarray iv ABSTRACT Ph.D THESIS IDENTIFICATION OF TISSUE SPECIFIC GENES BY COMPARATIVE ANALYSIS OF MICROARRAY DATA IN BOS TAURUS Selçuk KAPLAN THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY THE DEGREE OF DOCTOR OF ANIMAL SCIENCE IN AGRICULTURAL FACULTY Advisor: Prof. Dr. Saim BOZTEPE Year, 2013, 62 Jury Advisor Prof. Dr. Saim BOZTEPE Prof. Dr. Ramazan YETĠġĠR Prof. Dr. Murat Soner BALCIOGLU Prof. Dr. Birol DAĞ Doç. Dr. Fulya ÖZDĠL Tissue specific genes play important roles in development and metabolism. Currently, Gen Bank has 3.363.628 GEO Profiles and 397.988 microarray data related to various tissues. Although huge amounts of useful data are available to scientists in Gen Bank, there is a lack of a follow-up strategy to analysis and use these data to identify new tissue specific genes. This study suggest a unique new method to using and analyzing microarray and microarray related GEO Profiles to identify tissue specific genes. In this study, adipose tissue sellected as a target tissue in order to validate the method. First, the human and mouse microarray data were analyzed comparatively. To support the microarray data, adipose tissue related GEO Profiles were selected from Gen Bank. Subsequently, adipose tissue related microarray and GEO Profiles data were evaluated simultaneously. Finally, this method hypothesized a novel adipose specific gene called Chordin-like 1 (Chrdl1). In order to test the hypothesis, RT-PCR was performed for the bovine tissue distribution. According to the RT-PCR results, Chrdl1 was highly expressed adipose tissue versus other tissues (P<0.05). This result was support the hypothesis and demonstrated that a new method used in this study was useful in identification of new tissue specific genes. Keywords: Adipose tissue, Adipose specific gene, Atgl, Chrdl1, Fabp4, Microarray v ÖNSÖZ Akademik hayatıma baĢladığım günden beri, mesleki ve kiĢisel geliĢimimde büyük emeği olan ve tüm akademik çalıĢmalarımda ve doktora eğitimim süresince her koĢulda bilgi birikimini ve insani desteğini esirgemeyen değerli danıĢman hocam sayın Prof. Dr. Saim BOZTEPE’ye sonsuz teĢekkürlerimi sunuyorum. ÇalıĢmanın belkide en kritik noktasında materyalin seçimi ve temini aĢamasında yardımlarını esirgemeyen Dr. Ġbrahim AYTEKĠN, Uzman. Hüseyin BAYIR, Dr. Mustafa Orhun DAYAN, ArĢ. Gör. Yasin ALTAY ve değerli öğrenci arkadaĢlarıma, iĢletmelerini bize açan ve numunelerin alınmasına olanak sağlayan Yılet Entegre Tesislerinin değerli çalıĢanlarına ayrı ayrı teĢekkür ediyorum. Bununla birlikte, hayvan materyallerini bize sağlayan Selet Et Entegre Tesisleri adına Metin KÖRPE’ye ayrıca teĢekkürü bir borç bilirim. Doktora çalıĢmamın teorik kısmında yaptığı katkı, yönlendirme, teĢvik ve her konuda yardımları için Ohio State Üniversitesi Zootekni bölümünden Prof. Dr. Michael DAVIS’ e teĢekkürlerimi sunarım. ÇalıĢmam boyunca beni her zaman sabırla destekleyen aileme, doğumuyla beni dünyanın en mutlu insanı yapan biricik kızım Zeynep Naz KAPLAN’a ve çalıĢmanın baĢlangıcından sonuçlanmasına kadar bana her koĢulda destek olan sevgili eĢim Nuray KAPLAN’a bu çalıĢmayı atfederim. Selçuk KAPLAN KONYA-2013 vi ĠÇĠNDEKĠLER ÖZET .............................................................................................................................. iv ABSTRACT ..................................................................................................................... v ÖNSÖZ ........................................................................................................................... vi ĠÇĠNDEKĠLER ............................................................................................................. vii SĠMGELER VE KISALTMALAR .............................................................................. ix 1. GĠRĠġ ........................................................................................................................... 1 2. KAYNAK ARAġTIRMASI ....................................................................................... 4 2.1. Adipoz Doku .......................................................................................................... 4 2.2. Adipoz Spesifik Genler .......................................................................................... 7 2.2.1. Yağ asidi bağlayıcı protein (FABP4).............................................................. 7 2.2.2. Adipoz trigliserit lipaz (ATGL) ...................................................................... 8 2.2.3. Leptin .............................................................................................................. 9 2.2.4. Adipoz spesifik aday gen Chrdl1 (Chordin like-1) ....................................... 14 2.2.5. Farklı hayvan türlerinde mikroarray yöntemi kullanılarak yapılan çalıĢmalar ................................................................................................................................ 16 3. MATERYAL VE YÖNTEM.................................................................................... 18 3.1. Materyal ............................................................................................................... 18 3.1.1. Gen taraması ................................................................................................. 18 3.1.2. Doku örneklerinin alınması .......................................................................... 19 3.2. Yöntem................................................................................................................. 21 3.2.1. Havanların hazırlanması ............................................................................... 21 3.2.2. Sıvı azotla dokuların havanda ezilmesi ........................................................ 22 3.2.3. Dokulardan RNA izolasyonu ........................................................................ 22 3.2.4. RNA konsantrasyon saflıklarının belirlenmesi ............................................. 24 3.2.5. cDNA sentezi ................................................................................................ 29 3.2.6. Primer tasarımı .............................................................................................. 30 3.2.7. Real time pcr ................................................................................................. 30 3.2.8. Real time pcr sonuçlarının değerlendirilmesi ............................................... 32 3.2.9. Ġstatistik analizler .......................................................................................... 32 4. ARAġTIRMA SONUÇLARI VE TARTIġMA ...................................................... 33 4.1. Bilimsel hipotezin kurulması ............................................................................... 36 4.2. Hipotezin Real Time PCR ile Test Edilmesi ....................................................... 38 4.3. Yağ Asidi Bağlayıcı Protein 4 (FABP4).............................................................. 40 4.4. Adipoz Trigliserit Lipaz (ATGL) ........................................................................ 42 4.4. Chordin-like 1 (Chrdl1) ....................................................................................... 43 5. SONUÇLAR VE ÖNERĠLER ................................................................................. 44 vii 5.1. Sonuçlar ............................................................................................................... 44 5.2. Öneriler ................................................................................................................ 45 5.2.1. Ulusal karkas derecelendirme sisteminin kurulması .................................... 45 5.2.2. Markere dayalı seleksiyon yönteminin kullanılması .................................... 47 5.2.3. Sağlıklı beslenme konusunda toplumsal bilincin arttırılması ....................... 48 KAYNAKLAR .............................................................................................................. 51 ÖZGEÇMĠġ .................................................................................................................. 62 viii SĠMGELER VE KISALTMALAR DNA DNase RNA RNase cDNA mRNA dNTP A T G C bç GEO Profiles GEO DataSets g kg mg μg kDa μl ml °C dk sn SNP PCR RT-PCR rpm Chrdl1 FABP4 ATGL BMP HIF-1 TSE TNFα MCP-1 PAI-1 ASP LPL TSH FABPs Deoksiribonükleik asit Deoksi Ribo Nükleaz Ribonükleik asit Ribo Nükleaz Komplementer Deoksi Ribo Nükleik Asit Haberci RNA Deoksiribonükleozid trifosfat Adenin nükleotid Timin nükleotid Guanin nükleotid Sitozin nükleotid Baz çifti GeniĢ kapsamlı gen ekspresyon profilleri Gen ekspresyon veri setleri Gram Kilogram Miligram Mikrogram Kilo dalton Mikro litre Mili litre Santigrat derece Dakika Saniye Tek nükleotid polimorfizmi Polimeraz Zincir Reaksiyonu Ters transkriptaz Polimeraz Zincir Reaksiyonu (Rotate per minute) Dakikadaki devir sayısı Chordin-like 1 Yağ asidi bağlayıcı protein 4 Adipose trigliserit lipaz Bone morphogenetic protein Hypoxia inducible factor-1α Türk Standartları Enstitüsü Tumor necrosis factor-alpha Monocyte chemoattractant protein-1 Plasminogen activator inhibitor-1 Acylation stimulating protein Lipoprotein lipaz Thyroid stimulating hormone Yağ asidi bağlayıcı proteinler ix 1 1. GĠRĠġ Adipoz doku son yıllarda araĢtırmacılar tarafından vücut homeostazisinde önemli görevleri olan endokrin bir organ olarak gösterilmektedir (Galic ve ark., 2010). Adipoz dokuda meydana gelen problemler insanlarda obezite ve obeziteye bağlı metabolik rahatsızlıklara neden olurken çiftlik hayvanlarında ise yağlı karaciğer sendromu gibi metabolik bozukluklara neden olmaktadır (Umucalılar ve GülĢen, 2005; Maury ve Brichard, 2010). Bununla birlikte çiftlik hayvanlarında görülen aĢırı yağlanma, yenilebilir et miktarını azaltıp karkas randımanını olumsuz yönde etkilemek suretiyle ciddi ekonomik kayıplara neden olmaktadır (Anonim, 2013a). Söz konusu nedenlerden dolayı son yıllarda adipoz dokunun insanlarda ve çiftlik hayvanlarında vücut fonsiyonlarını ne yönde etkilediği büyük bir merak konusu olmuĢ; adipoz doku geliĢiminin moleküler mekanizmasının aydınlatılmasına yönelik çalıĢmalar hız kazanmıĢtır. Mikroarray analizleri adipoz dokunun geliĢiminden sorumlu doku spesifik gen ekspresyonlarının tespit edilmesinde kullanılan moleküler yöntemlerin baĢında gelmektedir. 1995 yılında ilk keĢfedilmesinden bu yana mikroarray teknolojileri, binlerce genin farklı biyolojik koĢullardaki gen ekspresyon profillerinin karĢılaĢtırılmasında baĢvurulan önemli bir moleküler yöntem olarak öne çıkmaktadır. (Schena ve ark., 1995). Günümüzde, yüksek hacimli mikroarray teknolojileri içinde gen ekspresyon çalıĢmaları, genotipleme, proteomiks, hücre biyolojisi, kanser biyolojisi, farmakogenomik, bulaĢıcı hastalıkların tanımlanması, besin örneklerinin içindeki patojen varlığının tespiti ve mikrobiyal izolatların tanımlanması gibi birçok uygulamaya ev sahipliği yapmaktadır (Kumar, 2009). Preadipozit hücrelerinin farklılaĢıp olgun adipozitlere dönüĢmesi sürecine adipogenesis denilmektedir (Anonymous, 2010). Adipogenesis transkripsiyon faktörleri ve hücre döngüsü proteinlerinin gen ekspresyonlarını regüle ederek adipozit geliĢimini sağladığı birden çok aĢaması olan bir prosestir (Lefterova ve Lazar, 2009). Adipoz geliĢiminin adipoz spesifik genler tarafından önemli derecede etkilendiği oldukça iyi bilinmektedir. Son yirmi yıldır, bazı memeli türlerinde insan, fare (Cornelius ve ark., 1994; Ntambi ve Cheul, 2000), domuz (Bai ve ark., 2008) ve sığırlarda (Fernyhough ve ark., 2007; Taniguchi ve ark., 2008) yağ hücrelerinden yağ sentezlenmesi üzerine doğrudan yada dolaylı olarak etkisi olduğu belirlenen bazı genler ortaya çıkarılmıĢtır. Söz konusu genlerin kodladığı peroksisome proliferator activated receptor γ (PPARγ) 2 ve CCAAT/enhancer-binding protein γ (C/EBPγ) gibi transkripsiyon faktörlerinin canlı ya da laboratuar ortamlarında yağ hücresi geliĢimini doğrudan etkilediği araĢtırmacılar tarafından ortaya konmuĢtur (Rosen ve ark., 2000; Fernyhough ve ark., 2007). Bu genler tarafından kodlanan ve genlerin ifadelerinin düzenlenmesinde görev alan transkripsiyon faktörleri genel olarak yağ ve yağ asidi metabolizmasında görev almaktadır. Bu faktörler tamamen bağımsız çalıĢmamaktadır, aksine bir çok önemli yolla iĢlevsel olarak birbirleriyle etkileĢim halinde görev yapmaktadır (Kim ve ark., 2006). Bu genler arasında PPARγ geni yağ hücrelerinin farklılaĢmasında görevli bir çok genin (örneğin; yağ depolama ve metabolizmasında görevli AP2 ve lipoprotein lipaz LPL gibi) ifadesini düzenleme görevini yürütmesinden dolayı yoğun olarak araĢtırmacıların ilgisini çekmektedir (Rosen ve ark., 2000; Fernyhough ve ark., 2007). Bütün bu geliĢmelere rağmen, yağ sentezinde merkezi görev alan transkripsiyon faktörleri ve bu faktörleri kodlayan genlerle ilgili açıklığa kavuĢmamıĢ birçok nokta bulunmaktadır. Bu durum söz konusu genlerle ilgili daha fazla araĢtırma yapılmasını gerektirmektedir. Günümüze kadar adipoz dokuda yüksek derecede ekspresyon gösterebilen 15 adipokin’in (adipoz spesifik genin) tespit edildiği gösterilmektedir (Piya ve ark., 2013). Adipoz geliĢiminin tamamen ortaya çıkarılması için çok daha fazla adipoz spesifik genin belirlenmesi ve bu genlerin adipoz dokunun geliĢimi üzerine etkilerinin ortaya konulması gerekmektedir. Yeni adipoz spesifik genlerin keĢfedilmesi sayesinde adipoz geliĢiminin genetik mekanizmasının ortaya çıkarılması, gerek insanlarda adipoz dokuya bağlı obezite, tip 2 diyabet ve kardiyovasküler hastalıkların tedavisinde gerekse çiftlik hayvanlarında üretim verimliliğinin arttırılmasında önemli rol oynayacaktır. Bu amaçla, adipoz spesifik genlerin bulunması ve fonksiyonlarının ortaya çıkarılması için yeni metotların geliĢtirilmesi büyük önem arz etmektedir. Günümüzde, gen bankasında adipoz doku ile ilgili olarak 1.346.912 GEO Profiles (GeniĢ kapsamlı gen ekspresyon profilleri) ve 13.324 GEO DataSets (Gen ekspresyon veri setleri) olmak üzere toplamda 1.360.236 mikroarray verisi bulunmaktadır (Anonymous, 2013a; Anonymous, 2013b). Gen bankasında adipoz doku ile ilgili bu kadar fazla mikroarray verisi olmasına rağmen bu verilerin karĢılaĢtırmalı analizi yapılarak yeni adipoz spesifik genlerin ortaya çıkrarılmasına yönelik herhangi bir çalıĢma bulunmamaktadır. Bu nedenle gerek mikroarray verilerinin karĢılaĢtırmalı analizine yönelik yeterli çalıĢma olmaması gerekse adipoz dokunun sahip olduğu 3 biyolojik önemden dolayı bu çalıĢmada, doku spesifik genlerin ortaya çıkarılması için geliĢtirilen yeni metodun uygulaması için adipoz doku hedef doku olarak belirlenmiĢtir. Tümüyle değerlendirildiğinde bu çalıĢmanın amacı, sığırlarda doku spesifik genlerin tespit edilmesine olanak sağlayan yeni bir metot geliĢtirmek ve söz konusu metodun bilimsel araĢtırmalarda kullanılabilirliğini ortaya koymaktır. 4 2. KAYNAK ARAġTIRMASI 2.1. Adipoz Doku Geleneksel olarak, adipoz doku yağların muhafaza edildiği bir depo olarak görülmekteydi. Günümüzde ise adipoz dokunun enerji metabolizmasında aktif rol oynayan, bazı hormon ve sitokinlerin kaynağı olarak tüm vücut metabolizmasında görev yapan bir doku olduğu belirtilmektedir. Adipozitlerin enerji metabolizmasındaki aktif rolü leptinin keĢfedilmesiyle ortaya çıkmıĢtır. Leptin hemen hemen tamamen adipozitlere özgü bir salgı ürünü olup hipotalamusta bulunan reseptörleri aracılığıyla hormonal fonksiyon göstermektedir (Flier, 1998). Bu bulgular adipoz dokunun pasif bir enerji deposu olduğu yönündeki düĢünceleri değiĢtirmiĢtir. Bu çalıĢmaların akabinde bilim adamları adipoz dokunun adipokin adı verilen bir çok biyoaktif peptidi salgıladığını ayrıca söz konusu adipokinlerin otokrin/parakrin ve sistemik (endokrin) düzeyde iĢlev gösterdiklerini bildirmiĢlerdir (Çizelge 2.1) (Kershaw ve Flier, 2004). Bunların ötesinde adipoz dokunun merkezi sinir sisteminde olduğu gibi hormonlardan gelen sinyallere karĢılık olarak bir çok reseptörü eksprese ettiği görülmektedir (Çizelge 2.2) (Kershaw ve Flier, 2004). Çizelge 2.1. Bazı adipozit türevi proteinler ve endokrin fonksiyonları (Kershaw ve Flier, 2004) Sitokinler ve sitokinlerle iliĢkili proteinler Diğer immun sistemle ilgili proteinler Fibrinolitik sistemle ilgili proteinler Komplement ve komplementle iliĢkili proteinler Leptin TNFα IL-6 MCP-1 PAI-1 Doku faktör Adipsin (komplement faktör D) Komplement faktör B ASP Lipidler ve lipid metabolizması ya da taĢınması ile ilgili proteinler Steroid metabolizmasıyla ilgili enzimler Adiponektin Lipoprotein lipaz (LPL) Kolesterol ester transfer protein Apolipoprotein E NEFA Stokrom P450 17βHSD 17βHSD1 5 Çizelge 2.2. Adipoz dokuda eksprese olan bazı reseptörler (Kershaw ve Flier, 2004) Geleneksel endokrin hormonları için reseptörler Ġnsulin reseptör Glucagon reseptör GH reseptör TSH reseptör Gastrin/CCK-B reseptör Glukagon benzeri peptid-1 reseptör Anjiyotensin 2 reseptörleri tip 1 ve tip 2 Nükleer hormon reseptörleri Glukokortikoid reseptör Vitamin D reseptör Tiroid hormon reseptör Androjen reseptör Estrojen reseptör Progesteron reseptör Leptin reseptör IL-6 reseptör TNFα reseptör Katekolamin reseptörleri β1, β2 ve β3 reseptörleri α1 ve α2 reseptörleri Adipoz dokuda meydana gelen anormal artıĢ ve azalmalar metabolik problemlere neden olmaktadır. Öyleki adipoz dokudaki anormal artıĢ obezite ve obeziteye bağlı insülin direnci, hiperglisemi, dislipidemi, kardiyovasküler ve hipertansiyon gibi önemli rahatsızlıklara neden olmaktadır (Grundy ve ark., 2004). Bununla birlikte deri altındaki adipoz dokudaki anormal azalmalar metabolizmanın bozulmasına, derinin hızla yaĢlanmasına ve sonuç olarak lipodistrofi rahatsızlığına neden olmaktadır (Anonim, 2013b). Adipoz doku memelilerde beyaz adipoz doku ve kahverengi adipoz doku olarak ikiye ayrılmaktadır (Cinti, 2005). Ruminantlarda doğumda adipoz dokunun büyük bir çoğunluğu kahverengi adipoz dokudan oluĢurken doğum sonrasında bu dokuların büyük bir çoğunluğu beyaz adipoz dokuya dönüĢmektedir. Yeni doğan buzağıların vücut ağırlığının % 1.5 – 2.0’si kahverengi adipoz dokudan oluĢmaktadır. Memelilerde beyaz adipoz doku böbreklere yakın bölgelerde, üreme organlarının çevresinde, perirenal depolarda, omental ve mezentrik depolarda ve kas içi ve kas aralarında bulunmaktadır (Mersmann ve Smith, 2004). Kahverengi adipoz doku ise memelilerde böbrek, kalp ve aort çevresinde interkostal kaslar ve sternum boyunca, boyunun iç kısımlarında ve ana arter ve damarların çevresinde görülmektedir (Stock ve Cinti, 2003). Ruminant hayvan karkaslarında adipoz doku derialtı, kasarası ve kas içi olarak yerleĢim göstermektedir (Öztürkcan ve ark., 1996). Beyaz adipozit hücrelerinden oluĢan beyaz adipoz doku adipoz dokunun büyük bir çoğunluğunu oluĢturmaktadır. Beyaz adipoz doku vücutta yerleĢim yerine bağlı 6 olarak viskeral (karın boĢluğu) ve subkutan (deri altı) olmak üzere isimlendirilmektedir (Avram ve ark., 2005). Beyaz adipoz doku, diyetten gelen fazla enerjiyi trigliserit olarak depolamaktadır. Açlık durumunda ise beyaz adipoz dokuda depo edilen trigliserit lipaz aktivitesiyle yağ asidi ve gliserole parçalanarak vücudun enerji ihtiyacı karĢılanmaktadır (Lieberman ve Marks, 2009). Bir yağ hücresinin çapı normalde 1-2 mikron arasında değiĢmesine rağmen, yağ biriktirdiğinde bu çap 120 mikrona kadar artabilmektedir. Bu durum yağ hücrelerinin yağ depolama kapasitesinin oldukça yüksek olduğunu açıkça göstermektedir (Öztürkcan ve ark., 1996). Beyaz adipoz doku ayrıca leptininde içinde bulunduğu biyoaktif peptid ve proteinleri (adipokinleri) sentezleyerek vücut homeostazisini sağlamada merkezi rol oynamaktadır (Millo, 2004). Kahverengi adipoz doku ise mitokondrice zengin bir dokudur. Kahverengi adipoz doku memelilerde vücut soğuğa maruz kaldığında oksidatif fosforilasyon yoluyla vücut sıcaklığını dengede tutmaktadır (Stock ve Cinti, 2003). Beyaz ve kahverengi adipoz dokunun elektron mikroskobu altındaki görüntüleri ġekil 2.1 ve 2.2’de gösterilmektedir (Anonim, 2013c). ġekil 2.1. Beyaz Adipoz Doku (Anonim, 2013c) 7 ġekil 2.2. Kahverengi Adipoz Doku (Anonim, 2013c) 2.2. Adipoz Spesifik Genler 2.2.1. Yağ asidi bağlayıcı protein (FABP4) FABPs (Yağ asidi bağlayıcı proteinler) ilk olarak 1972 yılında keĢfedilmiĢtir (Ockner ve ark., 1972). Yağ asidi bağlayıcı proteinler 14-15 kDa’luk proteinler olup 126-134 amino asitten oluĢmaktadır (Zimmerman ve Veerkamp, 2002). Yağ asidi bağlayıcı proteinler ile ilgili bir çok çalıĢma yapılmasına rağmen ilgili proteinlerin farklı dokulardaki fonksiyonları hala tam olarak anlaĢılamamıĢtır (Haunerland ve Spener, 2004). FABP4 (Yağ asidi bağlayıcı protein 4), yağ asidi bağlayıcı protein ailesinin en iyi tanımlanmıĢ üyesidir (Furuhashi ve HotamıĢlıgil, 2008). FABP4 geni hayvan türlerinde yoğun bir Ģekilde çalıĢılmaktadır. Cho ve ark. (2008), FABP4 geni polimorfizmlerinin Kore yerli sığır karkaslarında kabuk yağı kalınlığı üzerinde etkili olduğunu bildirmektedir. Gerbens ve ark. (1998), FABP4 geninin Duroc domuzlarında kas içi yağ birikimi üzerine etkileri olduğunu ifade etmektedir. Hoashi ve ark. (2008), Japon siyah sığırlarında FABP4 I74V genotipi ve kas içi yağ asidi kompozisyonu arasında önemli bir iliĢki olduğunu bildirmektedir (P<0.01). Siyah Alaca sığırlarda et kalitesi SCD (Sterol-Koenzim A desaturaz) ve FABP4 genotipleri tarafından etkilenmektedir (Narukami ve ark., 2011). 8 FABP4 geni, karkaslarında yüksek mermerizasyon gösteren Hanwoo sığırlarında düĢük mermerizasyon gösteren Hanwoo sığırlarına göre yüksek derecede ekspresyon göstermektedir (Lee ve ark., 2008). Xu ve ark. (2011), FABP4 AA genotipinin Çin yerli koyun ırklarında kas içi yağ (P<0.05), gevreklik (P<0.05) ve kas mermerizasyon skoru (P<0.05) ile önemli derecede iliĢki gösterdiğini bildirmektedir. Li ve ark. (2008), FABP4 geninin Beijingyou ve Jingxing piliçlerinde kas içi yağ yüzdesi üzerine önemli etkileri olduğunu ifade etmektedir. Bilim adamları tarafından yapılmıĢ olan çalıĢmalar, FABP4 geninin çiftlik hayvan karkaslarında mermerizasyon özelliğinin kontrol edilmesinde potansiyel bir hedef marker olduğunu göstermektedir. 2.2.2. Adipoz trigliserit lipaz (ATGL) Adipozitlerin lipolizinde 2004 yılına kadar sadece HSL (hormona duyarlı lipaz)’ın görev aldığı düĢünülmekteydi. Daha sonraları farklı bilim adamları tarafından yapılan çalıĢmalar HSL’ın olmadığı farelerin hala lipolitik aktivitelerini sürdürdüklerini göstermektedir. Bu durum akıllara trigliserit moleküllerini parçalayan farklı lipazların olabileceği düĢüncesini getirmiĢtir (Osuga ve ark., 2000; Wang ve ark., 2001; Holm, 2003). Daha sonra ATGL geninin trigliserit mobilizasyonunun ilk aĢamasında merkezi bir rol oynadığı araĢtırmacılar tarafından ortaya konulmuĢtur (Zimmermann ve ark., 2004) ATGL geniyle ilgili, hayvanlar üzerinde yapılan çalıĢmalara bakıldığında; Piliçlerde açlık durumunun ATGL gen ekspresyonunun hızla yükselmesine neden olduğu belirtilmektedir (Serr ve ark., 2009; Oh ve ark., 2011) Kosteli ve ark. (2010), farelerde kalori kısıtlamanın ATGL mRNA ekspresyonunu önemli derecede yükselttiğini bildirmektedir. Deiuliis ve ark. (2008), ATGL geninin kalori kısıtlaması yapılan domuzlarda yüksek derecede ekspresyon gösterdiğini ifade etmiĢlerdir. ATGL gen ekspresyonunun kuluçka sonrası açlık dönemindeki civcivlerin adipoz dokusunda artıĢ gösterdiği belirtilmektedir (Lee ve ark., 2009). Serr ve ark. (2011), ATGL gen ekspresyonunun, deksametazon enjeksiyonu yapılan kanatlılarda salin enjeksiyonu yapılanlara göre önemli derecede fazla olduğunu bildirmektedir. 9 Cui ve ark. (2011), ATGL geninin sığırlarda karkas ve et kalitesini belirlemede bir genetik marker olarak kullanabileceğini ifade etmektedir. Dai ve ark. (2011), domuzlardaki ATGL gen polimorfizmiyle (G/A392) iç organlardaki adipoz doku, yağsız et yüzdesi, kabuk yağı kalınlığı ve göz kası gibi karkas özellikleri arasında önemli bir iliĢki olduğunu bildirmektedir. Shan ve ark. (2008) domuzlarda ATGL gen ekspresyon düzeyinin 20 kg vücut ağırlığında (yaklaĢık 8 haftalık yaĢ) gözle görülür bir artıĢ gösterdiğini ve en yüksek seviyesine 40 kg vücut ağırlığında (yaklaĢık 12 haftalık yaĢ) ulaĢtığını bildirmektedir. ATGL geni ile ilgili olarak hayvanlarda yürütülen çalıĢmalar değerlendirildiğinde, kalori kısıtlamanın ATGL mRNA ekspresyonunu önemli derecede arttırdığı ve ATGL geninin karkas özellikleri için genetik marker olarak kullanılabileceği sonucuna varılmaktadır. 2.2.3. Leptin Friedman ve arkadaĢları ilk olarak 1994 yılında leptinin, obez (ob) geninin ürünü olan bir hormon olduğunu keĢfetmiĢlerdir (Castracane ve Henson, 2007). Leptinin keĢfi, uzun süreli besin alınımının regülasyonu, vücut ağırlığı, enerji harcama ve neroendokrin fonksiyonları gibi obezitenin oluĢumuna neden olan faktörlerin aydınlatılması açısından büyük önem taĢımaktadır (Millo, 2004). Leptin yapısal olarak sitokinlere benzerlik gösteren 16 kDa’luk bir polipeptid olup, 167 amino asitten oluĢmaktadır (Chan ve ark., 2000). Leptin özellikle farklılaĢma gösteren adipozitlerden eksprese olmaktadır. Subkutan adipoz doku leptin üretiminin % 80’nini üstlenmektedir. Laboratuvar koĢullarında yapılan hücre kültürü çalıĢmalarında, leptin üretiminin subkutan adipozitlerde diğerlerine oranla daha yüksek miktarda üretildiği görülmektedir (Puente ve ark., 2008). Leptinin öncelikli görevi vücut ağırlığı ve iĢtahı düzenlemektir. Leptin besin alınımını azaltmasının yanında, metabolik hızı ve termogenesisi yükselterek enerji tüketimini arttırmaktadır (Friedman ve Halaas, 1998; Webber, 2003). Leptinin enerji homeostazisinde gösterdiği fonksiyonlar oldukça iyi ortaya koyulmuĢtur. Bu fonksiyonların çoğu özellikle hipotalamik yollarla gerçekleĢmektedir. Diğer fonksiyonları ise kas ve pankreas hücreleri gibi perriferal dokular üzerinde gerçekleĢmektedir (Friedman ve Halaas, 1998; Bjorbaek ve Kahn, 2004). Leptin seviyesinin sirküle edilmesi, adipoz dokuda depolanan enerji miktarını ve doğrudan merkezi sinir sisteminin etkilenmesi sonucu enerji homeostazisi ve nöroendokrin 10 fonksiyonlarının düzenlenmesini etkilemektedir. Leptin hayvanlarda zayıflığa neden olmakta, leptinden yoksun hayvanlarda obezite görülmektedir (Falcão ve ark., 2012). Leptinin vücut ağırlığı homeostazisi üzerindeki rolü, 50 yıldan fazla bir süre önce farelerde kendiliğinden oluĢan obezite geni (ob) mutasyonu sonucu ortaya çıkmıĢ olup ob/ob genotipini taĢıyan fareler obezite gösterirken, OB/OB genotipini taĢıyan fareler normal vücut kondüsyonunda olmaktadır (Ingalls ve ark., 1950). ġekil 2.3’te normal ve obez fare bir arada gösterilmektedir (Anonymous, 2011a). ġekil 2.3. Normal ve obez fare (Anonymous, 2011a) Çiftlik hayvanları üreticileri için leptin ve yağlanma arasındaki iliĢki üretim verimliliği açısından büyük önem taĢımaktadır. Kandaki leptin hormonu konsantrasyonları yağlı hayvanlarda daha yüksek olmaktadır. Öyleki yağlı koyunların kanındaki leptin hormonu konsantrasyonunun yağsız koyunlardan 8 kat kadar fazla olduğu bildirilmektedir (Wylie ve Newforge, 2007). ġekil 2.4’te normal ve obez koyun birarada gösterilmektedir (Wylie ve Newforge, 2007). 11 ġekil 2.4. Normal ve obez koyun (Wylie ve Newforge, 2007) Buchanan ve ark. (2003), leptin geninin SNP (tek nükleotid polimorfizm) ile süt ve protein verimi arasındaki iliĢkileri araĢtırdıkları çalıĢmalarında, 416 adet Siyah Alaca sığırın genotiplemesini yapıp laktasyon performanslarının karĢılaĢtırılması için karıĢık model uygulaması yapmıĢlardır. AraĢtırma sonuçlarına göre, homozigot T alleline sahip hayvanların CC genotipine sahip hayvanlara göre günlük 1.5 kg daha fazla süt ürettiklerini ancak söz konusu farklılığın laktasyon süresi boyunca süt yağı ve süt protein yüzdesini etkilemediğini bildirmiĢlerdir. Chebel ve ark. (2008), leptin geninin R4C lokusu ekzon 2 bölgesinde SNP analizi ile Siyah Alaca sığırlarda laktasyon performası ve sağlık üzerine etkilerini araĢtırdıkları çalıĢmalarında, sığırlar günde 3 kez sağılmıĢ süt verimi ve süt komponentleri her hayvan için aylık olarak kayıt edilmiĢtir. Sığırlarda görülebilecek ayak sorunları, mastitis ve abomasum deplasmanı gibi rahatsızlıklar bireysel olarak takip edilmiĢtir. AraĢtırma sonuçlarına göre, CT genotipini taĢıyan hayvanlar ilk 62 günlük sağım periyodunda (2.98 ± 0.02) vücut kondüsyon skoru göstererek CC (3.02 ± 0.02) ve TT (3.04 ± 0.03) genotipini taĢıyan hayvanlara göre daha düĢük vücut kondüsyon skoru göstermiĢlerdir. Süt verimi ve komponentleri bakımından ise CC genotipi CT ve TT genotiplerine göre daha düĢük bir grafik göstermiĢtir. 305 günlük verime göre, % 3.5 yağa göre düzeltilmiĢ süt, süt yağı ve süt proteini bakımından CC 12 genotipli bireyler CT genotipli bireylere göre sırayla 258, 12 ve 10.7 kg daha az verime sahiptir. TT genotipli bireylerde abomasum deplansmanı bakımından % 4.3’ lük bir artıĢ görünmektedir. Clempson ve ark. (2011), Siyah Alaca sığırlarda leptin geninde SNP analizi ile üreme, büyüme ve süt verimi özellikleri arasındaki iliĢkileri araĢtırmıĢlardır. Elde edilen SNP’lerin erken kas geliĢimi ile iliĢkili olduğunu, A1457G ve A59V genotiplerinin sırasıyla kas boyu ve uzunluğuyla iliĢkili olduğu, UASMS1, UASMS2, A1457G ve A59V genotiplerinin üreme ile ve A59V genotipinin süt üretimiyle iliĢkili olduğunu tespit etmiĢlerdir. DeVuyst ve ark. (2008), besi sığırlarında doğum ağırlığı ve leptin SNP’leri arasındaki iliĢkileri araĢtırdıkları çalıĢmalarında CT ve TT genotipine sahip sığırların, CC genotiplerine sahip sığırlara göre daha yüksek doğum ağırlığına sahip olduklarını bildirmiĢlerdir. Komisarek ve ark. (2007), leptin geni polimorfizmleriyle Jersey sığırlarında üreme özellikleri arasındaki iliĢkileri araĢtırdıkları çalıĢmalarında leptin geninin R4C, A59V ve C(-963)T mutasyonlarını 219 sığırda PCR-RFLP yöntemi kullanarak test etmiĢlerdir. AraĢtırma sonuçlarına göre, allel frekansları R4C 0.80 (C) ve 0.20 (T), A59V 0.67 (C) ve 0.33 (T) ve C(-963)T 0.83 (C) ve 0.17(T) olarak bulmuĢlardır. Üreme özellikleriyle istatistik olarak önemli korelasyon gösteren sadece A59V mutasyonu olmuĢtur (P<0.05). A59V mutasyonunda TT genotipine sahip sığırların buzağılama aralığı ve servis periyodu, CT ve CC genotipini taĢıyan sığırlara göre daha kısa olmuĢtur. Aynı zamanda TT genotipine sahip sığırların buzağı baĢına tohumlama sayısı, CT ve CC genotipini taĢıyan sığırlara göre daha az olmaktadır. Kulig ve ark. (2009), Limuzin sığırlarında leptin geni polimorfizmleriyle büyüme özellikleri arasındaki iliĢkileri araĢtırmıĢlar. Toplam 129 Limuzin buzağıda, leptin geni polimorfizmlerininin vücut ağırlığı, günlük canlı ağırlık artıĢı, sağrı uzunluğu ve göğüs çevresi özellikleri üzerine etkilerini araĢtırmıĢlardır. A59V polimorfizminin 210 günlük yaĢta vücut ağırlığını önemli derecede etkilediğini (P<0.01) bildirmiĢlerdir. Bununla birlikte ayrıca A59V polimorfizminin 3 ile 210 günlük yaĢ arasında günlük canlı ağırlık artıĢını önemli derecede etkilediğini (P<0.05) tespit etmiĢlerdir. Lagonigro ve ark. (2003), sığırlarda yem tüketimiyle leptin genine ait 5 farklı SNP noktası arasındaki iliĢkileri araĢtırmıĢlardır. Sonuç olarak leptin geni ekzon 2 bölgesinde bulunan polimorfizmine ait olan AT genotipini taĢıyan hayvanların AA 13 genotipini taĢıyan hayvanlara göre ortalama olarak % 19 daha fazla yem tüketimine sahip olduklarını bildirmiĢlerdir. Liefers ve ark. (2003), leptin konsantrasyonları bakımından gebeliğin son döneminde ve laktasyon sırasında gözlenen dalgalanmaların, süt sığırlarında enerji dengesi, besin alımı, canlı ağırlık ve östrüs özellikleri üzerine etkilerini araĢtırmıĢlardır. Ġlkine gebe olan 304 Siyah Alaca sığırda ilk 80 günlük laktasyon süresince canlı ağırlık, kuru madde tüketimi, enerji dengesi ve süt verimiyle ilgili ölçümler yapmıĢlardır. Buzağılama sonrası 30 günle 80 gün arasındaki sürede leptin konsantrasyonlarının ölçülmesi amacıyla sığırlardan 2 haftada bir günün aynı saatinde sağım sonrası beslemeden önce kan örnekleri alınmıĢtır. AraĢtırmalarında leptin konsantrasyonlarının gebelik süresince yüksek olduğunu, ancak doğum sonrasında bir düĢme gerçekleĢtiğini, plazma leptin konsantrasyonlarının laktasyon sırasında negatif enerji tablosu oluĢtuğunda azaldığını, bu durumda leptin konsantrasyonlarının enerji dengesi üzerinde etkileri olabileceğini ve doğum sonrası leptin konsantrasyonlarının eski düzeyine gelmesinin negatif enerji tablosunun Ģiddetine bağlı olduğunu bildirmiĢlerdir. Besi sığırlarında büyüme özellikleri ve kabuk yağı ile leptin genine ait SNP verileri arasındaki iliĢkilerin araĢtırıldığı çalıĢmada toplam 1653 sığırda leptin genine ait UASMS2 ve R25C polimorfizmleri genotiplendirilmiĢtir. Besi baĢından besi sonuna kadar dört kez vücut ağırlıkları ölçülmüĢ ve kabuk yağı ölçümleri de ultrason kullanılarak yapılmıĢtır. AraĢtırma sonuçlarına göre, R25C polimorfizminin büyüme özellikleri üzerine önemli bir etkisi olmadığı, UASMS2 polimorfizminin canlı ağırlık üzerine önemli bir etkisi olduğu (P<0.001) belirlenmiĢtir. Bununla birlikte UASMS2 CC genotipinin besi baĢı ağırlığının en fazla olduğu; UASMS2 TT genotipli sığırların en hızlı canlı ağırlık artıĢına sahip olduğunu tespit etmiĢlerdir. Ultrasonla yapılan kabuk yağı ölçümlerinde her iki polimorfizm kombine olarak değerlendirilmiĢ olup R25CCC/UASMS2-TT genotipli sığırlarda kabuk yağının en az olduğu, R25C-CC/UASMS2TT genotipli sığırlar en hızlı kabuk yağı geliĢimine sahip olurken R25C-CC/UASMS2CC genotipli sığırlar en yavaĢ kabuk yağı geliĢimine sahip olduğu belirtilmektedir (Lusk, 2007). Schenkel ve ark. (2005), leptin geni tekli nükleotid polimorfizmleri ile karkas ve et kalitesi özellikleri arasındaki iliĢkileri toplam 1111 sığır üzerinde araĢtırmıĢlardır. Deneme sonuçlarına göre leptin geni E2JW ve E2FB polimorfizmlerinin yağ, yağsız et verimi ve yağlanma derecesi ve göz kası alanındaki etin gevreklik özellikleri üzerine sırasıyla (P<0.01, P<0.05 ve P<0.01) önem düzeyinde etkili olduğunu bildirmiĢlerdir. 14 TCAC, CCAT ve TTAC haplotiplerinin populasyonun % 88’ini oluĢturduğunu, CCTT haplotipinin diğer haplotiplere göre yağsız et ve yağlanma derecesi üzerine (P<0.01) önem düzeyinde daha etkili olduğunu; TTTT haplotipinin diğer haplotiplerden farklı olarak göz kası alanında görülen gevreklik üzerine (P<0.03) önem düzeyinde etki gösterdiğini ifade etmiĢlerdir. Da Silva ve ark. (2012), leptin ve leptin reseptör genlerinin tekli nükleotid polimorfizmlerinin büyüme ve ultrason karkas özellikleri üzerine olan iliĢkilerini 2162 Nellore sığırında incelemiĢlerdir. ÇalıĢmada E2FB markerinin ağırlık kazancı ve göz kası alanı ve kabuk yağı özelllikleriyle önemli iliĢkiler gösterdiği, leptin promotor bölgesi markerlerinden C963T ve UASMS1’in göz kası alanı ile önemli derecede iliĢkisi olduğunu ve T945M markerinin ağırlık kazancıyla iliĢkisi olduğunu bildirmiĢlerdir. Sonuç olarak araĢtırmacılar leptin ve leptin reseptör markerlerinin büyüme ve karkas özelliklerinin geliĢtirilmesi için markere dayalı seleksiyon kapsamında kullanılabileceklerini ifade etmiĢlerdir. Woronuk ve ark. (2012), besi sığırlarında kabuk yağı ve canlı ağırlık özellikleri ile leptin geninde sitozin ve timin nükleotidlerinin yer değiĢtirmesi (C/T) sonucu meydana gelen tekli nükleotid polimorfizmi arasındaki iliĢkileri araĢtırdıkları çalıĢmalarında kabuk yağı ve vücut ağırlığı ölçümleri için 136.286 sığırın genotiplendirilmesini yapmıĢlardır. AraĢtırma sonuçlarına göre, T allelinin kabuk yağı üzerinde önemli bir etkiye sahip olduğu (P<0.0001), TT, CT ve CC genotipli sığırların sırasıyla 6.79 ± 0.02, 6.49 ± 0.01 ve 6.28 ± 0.01 mm kabuk yağına sahip olduğunu belirlemiĢlerdir. Genotipin canlı ağırlık üzerine olan etkisi önemli bulunmuĢtur (P<0.0001). TT, CT ve CC genotipli sığırların sırasıyla 484.2 ± 0.7, 488.0 ± 0.5 ve 487.3 ± 0.6 kg canlı ağırlığa sahip olduklarını bildirmiĢlerdir. 2.2.4. Adipoz spesifik aday gen Chrdl1 (Chordin like-1) Chrdl1 sığırlarda X kromozomu üzerinde taĢınan 51.42 kDa’luk moleküler ağırlığa sahip 453 amino asitten oluĢan protein kodlayan bir gendir (Anonymous, 2013c). Chrdl1 geni neurogenesin-1, neuralin-1 ve ventroptin olarak da isimlendirilmektedir. Chrdl1 geni Chordin genine homoloji göstermektedir. Chordin geni Noggin genine benzer olarak BMP (bone morphogenetic protein) gen ailesi reseptörlerine bağlanarak aktivitelerini inhibe etmektedir (Piccolo ve ark., 1996; Zimmerman ve ark., 1996; Nakayama ve ark., 2001). AraĢtırmacılar Chrdl1 geninin 15 Chordin genine benzer bir özellik gösterip, BMP gen ailesinden BMP2, BMP4 ve BMP7 genlerinin aktivitesini inhibe ettiğini göstermektedir (Sakuta ve ark., 2001; Takahashi ve ark., 2003; Larman ve ark., 2009). Chrdl1 CR1, CR2 ve CR3 olarak isimlendirilen 3 tane sisteince zengin prokollojen tekrarlara sahip olup, CR1 ve CR3 BMP reseptörlerine bağlanmayı sağlamaktadır (Larrain ve ark., 2000). Sakuta ve ark. (2001), Chrdl1 geninin ventral retinada yüksek derecede ekspresyon gösterdiğini bildirmiĢlerdir. Nakayama ve ark. (2001) Chrdl1 geninin farelerde mezenkima dokusundan türev alan dermatom ve organ gözü mezenkiması, geliĢimin ileri evrelerinde geliĢim gösteren iskelet kondrositlerinde, böbrek tübüllerinde, gastrointestinal bölgelerde ve yetiĢkinlerde kemik stromal hücrelerinde yüksek derecede ekspresyon gösterdiğini bildirmektedir. Chrdl1 geninin farelerde aynı zamanda nöral katmanlarda, yumurtalık yüzeyi epitelinde, geliĢen kıl foliküllerinde, gut ve dorsal sinir düğümü köklerinde ekspresyon gösterdiği ifade edilmektedir (Coffinier ve ark., 2001; Bowen ve ark., 2009). Zheng ve ark. (2010), Neurogenesin 1 geninin nöral kök hücrelerini aktive ederek zarar gören nöronların tamirine katkıda bulunduğunu belirtmektedir. Larman ve ark. (2009), Chrdl1 ve BMP7 genlerinin birbirine zıt hareket ettiğini bildirmektedir. Sağlıklı proksimal tübüllerde Chrdl1 geni BMP7 geninin ekspresyonunu baskılayarak kendi gen ekspresyonunu arttırmaktadır. BMP7 geni epitel hücrelerinin yeniden farklılaĢmasında görev almaktadır. Hasar gören böbreğin iyileĢmesi için epitel doku Chrdl1 gen ekspresyonunu azaltıp BMP7 gen ekspresyonunu arttırmaktadır. Kane ve ark. (2008), HIF-1 (Hypoxia inducible factor-1α) geninin Chrdl1 gen ekspresyonunu düzenlediğini bildirmektedir. Oksijen eksikliği görülen ortamlarda (hypoxia) Chrdl1 gen ekspresyonunun insan retina perisitlerinde artıĢ gösterdiği ifade edilmektedir. Bunun akabinde BMP4 gen sinyalini baskılayarak retinal anjiyogenesise katkıda bulunduğu belirtilmektedir. Chrdl1 geni ile ilgili literatür bildiriĢleri değerlendirildiğinde, Chrdl1 geninin genel olarak BMP gen ailesiyle iliĢki halinde olduğu ve BMP gen ailesinin fonksiyonlarının düzenlenmesinde merkezi rol oynadığı sonucuna varılmaktadır. 16 2.2.5. Farklı hayvan türlerinde mikroarray yöntemi kullanılarak yapılan çalıĢmalar Li ve ark. (2009), fare dokularında yağ geliĢimini etkileyen genleri araĢtırdıkları çalıĢmalarında, ISG12b1 geninin adipoz dokuda diğer dokulara göre önemli derecede fazla eksprese olduğunu bulmuĢlardır. AraĢtırmacılar, söz konusu genin ekspresyonu ile dokudaki yağ geliĢimi arasında negatif bir iliĢkinin olduğunu ortaya koymuĢlardır. ÇalıĢmada, soğuğa maruz bırakılan farelerin dokularında yağ üretimi artarken genin ekspresyon seviyesinin hızla düĢtüğünü tespit etmiĢlerdir. Çevre Ģartları normale döndüğünde ise genin ekspresyon seviyesinin yükseldiği ve yağ üretimini baskı altına alarak durdurduğunu belirlemiĢlerdir. Zhang ve ark. (2010), Qinchuan sığırlarında ette gevreklik üzerine etkili genleri araĢtırdıkları çalıĢmalarında, diĢi ve erkek Qinchuan sığırlarında göz kası alanı üzerinden alınan doku örneklerinde Sığır Genom Arrayleri kullanarak farklı düzeylerde eksprese olan genleri tespit etmeye çalıĢmıĢlardır. AraĢtırma bulgularına göre arraylerde bakılan 10.000 gen arasında 598 genin farklı düzeylerde eksprese olduğu tespit edilmiĢtir. Bu genlerin öncelikli olarak hücre adhezyonu, kollejen fibril organizasyonu ve sentezi, immun cevaplarda ve hücre matrix adhezyonunda görev aldığı belirtilmektedir. Yarru (2008), kanatlılarda aflotoksinli yem tüketiminin gen ekspresyon seviyelerinde yaptığı değiĢiklikleri araĢtırdıkları çalıĢmasında, gruplardan birine 2 mg AF/kg içeren yem vermiĢtir. Yapılan mikroarray analizi sonucunda aflotoksinli yem tüketen grupla kontrol grubu arasında 177 genin farklı düzeyde eksprese olduğunu tespit etmiĢtir. Bu genler arasında 97 genin yüksek düzeyde eksprese olduğu 80 genin ise düĢük seviyede ekprese olduğunu ortaya koymuĢtur. Hou ve ark. (2010), sığır meme dokusunda laktasyon sırasında farklı seviyelerde eksprese olan genleri mikroarray yöntemi kullanarak araĢtırdıkları çalıĢmalarında, laktasyon sırasında toplam 122 genin farklı seviyede ekspresyon gösterdiğini belirlemiĢlerdir. Ayrıca laktasyondaki hayvanlarla düveler ve kuru dönemdeki hayvanlar kıyaslandığında belirlenen 122 genden 79’unun yüksek düzeyde eksprese olduğu 43’ünün ise düĢük seviyede ekprese olduğunu ortaya koymuĢlardır. Wang ve ark. (2005), Siyah Alaca ve Japon Siyah sığırlarında göz kası dokusunda gen ekspresyon farklılıklarını araĢtırdıkları çalıĢmalarında, 3 Siyah Alaca ve 3 Japon Siyah sığırının göz kası dokusundan biyopsiyle örnek almıĢlardır. Daha sonra 17 alınan örnekler üzerinde mikroarray teknolojisi kullanarak gen ekspresyon seviyelerini araĢtırmıĢlardır. AraĢtırma bulgularına göre göz kası doku örneklerinde Japon Siyah sığırlarında 17 genin, Siyah Alaca sığırlarında ise 7 genin yüksek düzeyde eksprese olduğu tespit edilmiĢtir. Alınan sonuçları doğrulamak için Japon Siyah sığırlarında tespit edilen 17 gen içinden 6 gene kantitatif Real Time PCR uygulaması yapılmıĢtır. Sonuç olarak bu genlerin Japon Siyah sığırlarında yağ asidi sentezi ve yağ depolanmasında görev alabileceğini ifade etmiĢlerdir. Yamasaki ve ark. (2006) yaptıkları çalıĢmada, daha önce sığır kas içi yağ dokusunda tespit etkileri pGPx geninin yağ üretimi üzerine olan etkilerini mikroarray teknolojisi kullanarak araĢtırmıĢlardır. Sonuç olarak yağ sentezi sırasında transkripsiyonel faktör C/EBPδ ve pGPx geninin artıĢ gösterdiğini belirlemiĢlerdir. Bununla birlikte pGPx geninin yağ sentezi sırasında böbrek ve karın boĢluğu yağ dokusunda yoğun bir Ģekilde ekspresyon gösterdiğini tespit etmiĢlerdir. Clark, (2008). sığır etlerinde mozaikleĢmeyi arttıran genleri mikroarray teknolojisi kullanarak araĢtırdığı çalıĢmasında, kesilen 28 sığırdan göz kası doku örnekleri almıĢtır. Daha sonra bu örnekleri mozaikleĢme derecesine göre iki gruba ayırmıĢtır. Örnekler üzerinde ayrı ayrı yürütülen mikroarray analizleri sonucunda 9 genin farklı düzeylerde ekspresyon gösterdiğini tespit etmiĢtir. MozaikleĢme oranı fazla olan örneklerde söz konusu 9 gen arasında bulunan MYH3, HOXD10, MXRA8 ve CASQ2 genlerinin ekpresyonunun yüksek olduğu tespit edilmiĢtir. Bununla birlikte mozaikleĢme oranı fazla olan örneklerde NPNT, MRC1, DNER ve CYPB4 genlerinin düĢük seviyede ekspresyon gösterdiği belirtilmektedir. Geriye kalan ACTN2 geninin fonksiyonunun tam olarak anlaĢılamadığını bildirmiĢlerdir. 18 3. MATERYAL VE YÖNTEM 3.1. Materyal 3.1.1. Gen taraması Bu çalıĢmanın ana hedefini sığır dokularında daha önce literatürde bildiriĢi olmayan adipoz spesifik aday genlerin tespiti oluĢturmaktadır. Söz konusu aday genlerin tespiti konusunda gen bankasında derinlemesine analizler yapılmıĢtır. Aday genlerle ilgili sonuçların desteklenmesi amacıyla çeĢitli biyoinformatik ve istatistik programlarından yararlanılmıĢtır. Adipoz spesifik genleri araĢtırmak için gen bankasında bulunan GEO veritabanında “adipocyte” ve “adipose tissue” anahtar kelimeleri kullanılarak tarama yapılmıĢtır (Anonymous, 2013a; Anonymous, 2013b). Yapılan taramalar neticesinde GEO veritabanında söz konusu anahtar kelimelerle ilgili olarak farelerde GDS3142 various tissues (Mus musculus) ve insanlarda GDS596 Large-scale analysis of the human transcriptome (HG-U133A) mikroarray verilerine ulaĢılmıĢtır. GDS3142 mikroarray verisi 10 – 12 haftalık yaĢtaki farelerden alınan 22 farklı dokudan elde edilmiĢtir (Thorrez ve ark., 2008). GDS596 mikroarray verisi ise farklı kaynaklardan alınan fizyolojik olarak normal olan 79 insan dokusundan elde edilmiĢtir (Su ve ark., 2004). Her bir mikroarray ile ilgili veri seti GEO veritabanından indirilmiĢtir. GDS3142 veri seti dosyası farklı dokulardaki gen ekspresyon profillerinden oluĢmaktadır. 22 farklı doku içerisinden kas dokusu hariç diğer dokulardan 3 farklı biyolojik tekerrür bulunmaktadır. Kas dokusu ise 4 farklı biyolojik tekerrürden oluĢmaktadır. Aday genlerin belirlenmesinde kullanılacak yeni veri setini oluĢturmak amacıyla 7 farklı ana dokunun biyolojik tekerrürü ile ilgili veriler; dalak (GSM252067, GSM252068, GSM252069), kas (GSM252070, GSM252071, GSM252072, GSM252073), adipoz (GSM252093, GSM252094, GSM252115), akciğer GSM252095), (GSM252080, kalp (GSM252113, GSM252081, GSM252114, GSM252082), karaciğer (GSM252074, GSM252075, GSM252076) ve böbrek (GSM252083, GSM252084, GSM252085) ana veri setinden süzülerek yeni bir Excel dosyası formüle edilmiĢtir. Daha sonra dokularla ilgili gen ekspresyonlarının ortalaması alınmıĢtır. Adipoz spesifik genlerin tespiti için adipoz dokunun gen ekspresyonu diğer dokuların gen ekspresyon değerlerine bölünmüĢtür. Dokularla ilgili ortak bir katsayının ortaya çıkarılması için 19 elde edilen verilerin yeniden ortalaması alınmıĢtır. Sonuç olarak diğer dokuların gen ekspresyon değerlerinin adipoz dokuya göre oranını gösteren ortak bir katsayı bulunmuĢtur. GDS596 mikroarray ana veri setini oluĢturan 79 dokunun her biri 2 biyolojik tekerrürden oluĢmaktadır. Aday genlerin belirlenmesinde kullanılacak yeni veri setini oluĢturmak GSM18950), amacıyla karaciğer kalp (GSM18951, (GSM18953, GSM18952), GSM18954) akciğer adipozit (GSM18949, (adipoz hücresi) (GSM18975, GSM18976), iskelet kası (GSM19013, GSM19014), ve böbrek (GSM18955, GSM18956) ile ilgili veriler ana veri setinden süzülerek yeni bir Excel dosyası formüle edilmiĢtir. Daha sonra GDS3142 mikroarray veri setine uygulanan iĢlemlerin benzeri bir iĢlem GDS596 mikroarray veri setine uygulanarak diğer dokuların gen ekspresyon değerlerinin adipoz hücresine göre oranını gösteren ortak bir katsayı bulunmuĢtur. 3.1.2. Doku örneklerinin alınması Doku örneklerinin alınması aĢaması Konya’nın Sarayönü Ġlçesine bağlı Ladik Kasabasında bulunan Yılet Et Entegre Tesisleri'nde gerçekleĢtirilmiĢtir (ġekil 3.1). ġekil 3.1. Yılet Entegre Tesisinden bir görünüm (Anonim, 2012a) 20 ġekil 3.2. Angus Sığırları (Anonymous, 2012a) ÇalıĢmada kullanılan hayvansal doku örnekleri, yapılan görüĢmeler sonucu Selet et entegre tesislerinden gelen 5 farklı Angus sığırın yağ, kalp, karaciğer, dalak, böbrek, kas ve akciğer olmak üzere 7 farklı doku bölgesinden alınmıĢtır. ġekil 3.2’de Angus sığırları gösterilmektedir. ÇalıĢmada doku spesifik aday genlerin belirlenmesi amaçlandığı için dokuların birbirine karıĢma olasılığının ortadan kaldırılması gerekmektedir. Bu amaçla, dokuların alınması sırasında kontaminasyonu önlemek amacıyla doku konusunda uzman personelden yardım alınmıĢtır. ġekil 3.3’de kesim sonrasında angus sığırlarından alınan yağ, kalp, karaciğer, akciğer, dalak, böbrek ve kas dokuları gösterilmektedir. Alınan dokular derhal küçük parçalara ayrılıp, numaralandırılarak DNase-RNase free mikrosantrifüj tüplerine konmuĢtur. RNA tek zincirli yapıya sahip olduğu için DNA’ya göre çok daha hızlı degrede olmaktadır. Bunun yanında ortamda bulunan Ribo Nükleazlar RNA’nın degrede olmasına katkı sağlamaktadır. Bu nedenle olası RNA kayıplarını önlemek amacıyla dokular derhal kuru buz içine koyularak soğuk zincire alınmıĢtır. Laboratuvara getirilen doku örnekleri gerekli kontroller yapıldıktan sonra RNA izolasyonu yapılıncaya kadar -80 °C’ de tutulmuĢtur. 21 ġekil 3.3. Sığır doku örnekleri (Yağ, kalp, karaciğer, akciğer, dalak, böbrek ve kas) 3.2. Yöntem 3.2.1. Havanların hazırlanması RNA izolasyonunda kullanılan havanlar ilgili solusyonlar kullanılarak özel olarak dezenfekte edilmektedir. Dokuların havanda ezilmesi sırasında RNA degredasyonunu engellemek için havanlar önceden alüminyum folyolere sarılarak 24 saat boyunca -80 °C’lik buzdolabında muhafaza edilmektedir. ġekil 3.4’te çalıĢmada kullanılan porselen havan gösterilmektedir. ġekil 3.4. Porselen havan (Anonim, 2011a) 22 3.2.2. Sıvı azotla dokuların havanda ezilmesi Dokuların RNA izolasyonu sırasında homojen olarak çözünebilirliğini arttırmak amacıyla tüplerden alınan doku örnekleri steril havan içinde sıvı azotta ezilip toz haline getirilerek bir ön iĢleme tabi tutulmuĢtur. ġekil 3.5. Sıvı azotun tanka doldurulması (Anonim, 2010a) 3.2.3. Dokulardan RNA izolasyonu RNA izolasyonu için -80 °C’ de tutulan sığır doku örnekleri soğuk zincirde laboratuvara getirilmiĢtir. Sıvı azotta ezilmek suretiyle ön iĢleme tabi tutulan doku örneklerinden GF-1 Total RNA extraction kit (Vivantis) kiti kullanılarak RNA izolasyonu iĢlemi yapılmıĢtır. Ġzolasyonda 2 farklı protokol kullanılmıĢtır. 1. Protokol yağ, karaciğer, akciğer, böbrek ve dalak dokuları için kullanılmıĢtır. 2. Protokol RNA izolasyonunun daha zor olduğu fibröz (kalp ve kas) dokularında kullanılmıĢtır. Ġzolasyon aĢamaları aĢağıda verilmiĢtir. 1. Protokol a) Hassas terazide 30 mg hayvansal doku tartılmıĢtır. b) Tartılan doku derhal sıvı azotta ezilerek toz haline getirilmiĢtir. 23 c) Toz haline gelen doku örneği 2 ml’lik steril santrifüj tüpüne alındıktan sonra üzerine 700 μl Buffer TR çözeltisi (2-merkaptetanol önceden ilave edilmiĢ) eklenerek el mikseri yardımıyla homojenizasyon yapılmıĢtır. d) Örnekler maksimum hızda 3 dk santrifüj edilmiĢtir. e) Tüplerin üzerinde biriken üst faz dikkatlice alınarak filtreli homojenizasyon tüplerine aktarılmıĢtır. f) Örnekler maksimum hızda 2 dk santrifüj edilmiĢtir. g) Filtre alınarak aĢağıda biriken sıvı kısmın üzerine 650 μl 80%’ lik etanol ilave edilmiĢtir. Pipetleme yoluyla karıĢtırma iĢlemi yapılmıĢtır. h) OluĢan karıĢımın hepsi filtreli RNA bağlama tüplerine aktarılmıĢtır. i) Örnekler 10.000 x g hızda 1 dk santrifüj edilmiĢtir. j) Örneklerin üzerine 500 μl Wash Buffer çözeltisi ilave edilerek, 1 dk maksimum hızda santrifüj edilmiĢtir. k) RNA bağlama tüpünün üzerindeki filtre kaldırılarak aĢağıda biriken sıvı kısım dökülmüĢtür. l) RNA bağlama tüpünün içindeki membranın merkezine 70 μl DNase I Digestion Mix eklenmiĢ 15 dk oda sıcaklığında inkübe edilmiĢtir. m) Örneklerin üzerine 500 μl Inhibitor Removal Buffer eklenip 1 dk maksimum hızda santrifüj edilmiĢtir. Altta kalan sıvı kısım dökülmüĢtür. n) Örneklerin üzerine 500 μl Wash Buffer çözeltisi ilave edilerek 10.000 x g hızda, 1 dk santrifüj edilmiĢtir. Altta kalan sıvı kısım dökülmüĢtür. o) Örneklerin üzerine 500 μl Wash Buffer çözeltisi ilave edilerek 10.000 x g hızda, 1 dk santrifüj edilmiĢtir. Altta kalan sıvı kısım dökülmüĢtür. p) Örnekler 10.000 x g hızda, 1 dk santrifüj edilmiĢtir. r) RNA bağlama tüpü 1.5 ml’lik yeni bir steril santrifüj tüpüne aktarılmıĢtır. Örneklerin üzerine 60 μl RNase-free Water ilave edilerek, 1 dk inkübe edilmiĢtir. Daha sonra örnekler 10.000 x g hızda 1 dk santrifüj edilmiĢtir. s) Elde edilen RNA’lar kullanım zamanına kadar -80 °C’ de muhafaza edilmiĢtir. 2. Protokol a) Hassas terazide 30 mg hayvansal doku tartılmıĢtır. b) Tartılan doku derhal sıvı azotta ezilerek toz haline getirilmiĢtir. 24 c) Toz haline gelen doku örneği 2 ml’lik steril santrifüj tüpüne alındıktan sonra üzerine 300 μl Buffer TR çözeltisi (2-merkaptetanol önceden ilave edilmiĢ) eklenerek el mikseri yardımıyla homojenizasyon yapılmıĢtır. d) OluĢan lizat üzerine 590 μl su ilave edilmiĢ sonrasında da 10 μl Proteinaz K çözeltisi ilave edilmiĢtir. Vorteks yoluyla karıĢtırma yapılmıĢtır. Örnekler 65 °C’ de 10 dk inkübe edilmiĢtir. e) Örnekler 10.000 rpm hızda 3 dk santrifüj edilmiĢtir. f) Lizat filtreli homojenizasyon tüplerine aktarılmıĢtır g) 450 μl saf etanol ekleyip pipetleme yoluyla karıĢım sağlanmıĢtır. h) OluĢan karıĢımın hepsi filtreli RNA bağlama tüplerine aktarılmıĢtır. i) Örnekler 10.000 x g hızda 1 dk santrifüj edilmiĢtir. j) Örneklerin üzerine 500 μl Wash Buffer çözeltisi ilave edilerek, 1 dk 10.000 rpm hızda santrifüj edilmiĢtir. k) RNA bağlama tüpünün üzerindeki filtre kaldırılarak aĢağıda biriken sıvı kısım dökülmüĢtür. l) RNA bağlama tüpünün içindeki membranın merkezine 70 μl DNase I Digestion Mix eklenmiĢ 15 dk oda sıcaklığında inkübe edilmiĢtir. m) Örneklerin üzerine 500 μl Inhibitor Removal Buffer eklenip, 1 dk 10.000 rpm hızda santrifüj edilmiĢtir. Altta kalan sıvı kısım dökülmüĢtür. n) Örneklerin üzerine 500 μl Wash Buffer çözeltisi ilave edilerek 10.000 x g hızda, 1 dk santrifüj edilmiĢtir. Altta kalan sıvı kısım dökülmüĢtür. o) Örneklerin üzerine 500 μl Wash Buffer çözeltisi ilave edilerek 10.000 x g hızda, 1 dk santrifüj edilmiĢtir. Altta kalan sıvı kısım dökülmüĢtür. p) Örnekler 10.000 x g hızda, 1 dk santrifüj edilmiĢtir. r) RNA bağlama tüpü 1.5 ml’lik yeni bir steril santrifüj tüpüne aktarılmıĢtır. Örneklerin üzerine 60 μl RNase-free Water ilave edilerek 1 dk inkübe edilmiĢtir. Daha sonra örnekler 10.000 x g hızda, 1 dk santrifüj edilmiĢtir. s) Elde edilen RNA’lar kullanım zamanına kadar -80 °C’ de muhafaza edilmiĢtir. 3.2.4. RNA konsantrasyon saflıklarının belirlenmesi Ġzolasyonu tamamlanan RNA örneklerinin saflık ve miktarının belirlenmesi amacıyla NanoDrop Spektrofotometre ND 1000 cihazıyla ölçümler yapılmıĢtır. 260 ve 280 nm dalga boyları genel olarak RNA saflığının belirlenmesinde kullanılan 25 parametrelerdir. 260 ve 280 nm dalga boylarının birbirine oranlanmasıyla elde edilen katsayının ~ 2.0 civarında olması elde edilen RNA’nın saf olduğunu göstermektedir. Bunun aksi durumunda kontaminasyon olduğu kabul edilmektedir (Anonymous, 2011b). Bazı doku örneklerinde yapılan NanoDrop ölçümlerinden elde edilen diyagramlar gösterilmektedir (ġekil 3.6, 3.7, 3.8, 3.9, 3.10, 3.11, 3.12). ġekil 3.6. Akciğer Dokusu NanoDrop Ölçümü 26 ġekil 3.7. Böbrek Dokusu NanoDrop Ölçümü ġekil 3.8. Dalak Dokusu NanoDrop Ölçümü 27 ġekil 3.9. Kalp Dokusu NanoDrop Ölçümü ġekil 3.10. Karaciğer Dokusu NanoDrop Ölçümü 28 ġekil 3.11. Kas Dokusu NanoDrop Ölçümü ġekil 3.12. Yağ Dokusu NanoDrop Ölçümü 29 3.2.5. cDNA sentezi cDNA sentezi 1 μg total RNA’dan 2-steps RT-PCR Kit (Vivantis) kiti kullanılarak gerçekleĢtirilmiĢtir. cDNA sentezinde kullanılan karıĢımlar ve protokol aĢağıda gösterilmektedir. cDNA protokolü a) RNA primer karıĢımı 0.2 ml’lik mikrosantrifüj tüplerine hazırlanmaktadır. KarıĢım Çizelge 3.1’de gösterilmektedir. Çizelge 3.1. RNA primer karıĢımı BileĢenler Miktar Total RNA 1 μg Oligo d(T) (40μM) 1μl 10mM dNTPs 1μl Su ile hacim 10 μl’ye tamamlanır b) KarıĢım hazırlandıktan sonra örnekler 65 °C’ de 5 dk inkübe edilir ve buz üzerinde 2 dk bekletilir. c) Örnekler kısa süre santrifüj edilir. cDNA sentez aĢamasına geçilir. cDNA sentez karıĢımı Çizelge 3.2’de gösterilmektedir. Çizelge 3.2. cDNA sentez karıĢımı BileĢenler Miktar 10X Buffer M-MuLV 2 μl M-MuLV Reverse Transcriptase 100 unit Su ile hacim 10 μl’ye tamamlanır d) Her bir RNA primer karıĢımına cDNA sentez karıĢımından 10 μl ilave edilmektedir. e) Örnekler dikkatlice karıĢtırılıp 1 dk santrifüj yapılmaktadır. f) PCR’da 42 °C’ de 60 dk inkübasyon. g) Reaksiyon 85 °C’ de 5 dk inkübasyon ile sonlandırılıp örnekler 1 dk buz üzerinde bekletilmektedir. Sentezlenen cDNA direkt olarak PCR reaksiyonlarında kullanılabilmektedir. 30 3.2.6. Primer tasarımı Primer tasarımı için, çalıĢmaya konu olan Chordin-like 1 (Chrdl1) ve çalıĢmada housekeeping gen olarak kullanılan Cyclophilin genleriyle ilgili olarak NCBI (National Center for Biotechnology Information) Gen Bankası’ndan sığırlara ait sekanslar kullanılmıĢtır. AraĢtırma sonuçlarına göre Chordin-like 1 Bos taurus (Gene ID: 540275) ve Cyclophilin Bos taurus (Gene ID: 281419) sekansları primer tasarımı için tespit edilmiĢtir (Anonymous, 2013c). Primerlerin seçimi, sekansın baĢlat ve bitiĢ kodonları arasındaki sekanstan yapılmıĢtır. Primer tasarımında Tm Calculator (Anonymous, 2012b) ve Vector NTI 5.0 (Anonymous, 2012c) programlarından yararlanılmıĢtır. Chordin-like 1 (Chrdl1) ve Cyclophilin genleriyle ilgili tasarlanan primerler Çizelge 3.3’de gösterilmektedir. Çizelge 3.3. Kullanılan PCR primerleri Primer adı Nükleotid dizisi Erime sıcaklığı (°C) GC oranı (%) Uzunluk (bç) Chrdl1 F1 Chrdl1 R1 5`- ACTCCATCACTTCAAGCTGGTGA -3` 61,91 48 23 5`- TGCAGTCCAGCTGCAGCTT -3` 61,86 58 19 Cyclophilin F1 5`- TTCCATCGTGTGATCAAGGA -3` 58,43 45 20 Cyclophilin R1 5`- TTAAGCTTGAAGTTCTCATCGG -3` 57,24 41 22 3.2.7. Real time pcr cDNA’ ların Real Time PCR ile çoğaltımında Qiagen firmasına ait Quantitect SYBR PCR Mix kullanılmıĢtır. Aynı PCR Ģartlarına farklı kuyucuklarda Cyclophilin ve Chordin-like 1 (Chrdl1) master mixleri hazırlanarak ayrı kuyucuklarda denemeler gerçekleĢtirilmiĢtir. Cyclophilin ve Chordin-like 1 (Chrdl1) genleri için hazırlanan master mixler Çizelge 3.4. ve Çizelge 3.5’de gösterilmektedir. Çizelge 3.4. Chordin-like 1 (Chrdl1) master mix BileĢenler Miktar SYBR PCR Mix 12.5 μl Forward primer (5 μlM) 3 μl Reverse primer (5 μlM) 3 μl cDNA 2 μl dH2O 4.5 μl TOPLAM 25 μl 31 Çizelge 3.5. Cyclophilin master mix BileĢenler Miktar SYBR PCR Mix 12.5 μl Forward primer (5 μlM) 3 μl Reverse primer (5 μlM) 3 μl cDNA 2 μl dH2O 4.5 μl TOPLAM 25 μl Real Time PCR denemesi (Fluorion Real-Time PCR Iontek, Turkey) cihazında gerçekleĢtirilmiĢtir. PCR programı; 95 °C’de 15 dk sonrasında 50 döngü, 95 °C’de 30 s, 60 °C’de 35 s, 72 °C’de 40 s (Bu aĢamada floresan ölçüldü) ve son olarak da 60 °C’den 99 °C’ye kadar 0.5 derecelik artıĢla 99 °C’ye kadar çıkılarak erime sıcaklığı ölçülmüĢtür. ġekil 3.13’te Fluorion Real-Time PCR Iontek cihazı gösterilmektedir. ġekil 3.13. Fluorion Real-Time PCR Iontek 32 3.2.8. Real time pcr sonuçlarının değerlendirilmesi Real Time PCR sonuçlarının değerlendirilmesinde 2∆∆Ct Livak metodu kullanılmıĢtır (Livak ve Schmittgen, 2001). Metodun uygulanması için validasyon deneyleri dizayn edilmiĢtir. Validasyon deneyi için RNA izolasyonundan elde edilen cDNA’lar 10 kat, 100 kat ve 1000 kat olmak üzere 3 log sulandırılmıĢtır. Cyclophilin ve Chordin-like 1 (Chrdl1) mixleri ile her bir sulandırma üçer kez çalıĢılmıĢtır. Daha sonra gen ekspresyonlarının belirlenmesinde aĢağıdaki aĢamalar izlenmiĢtir. ΔCtyağ = Ctchordin - Ctcyclophilin bu değer, yağ dokusu için 2.28 bulunmuĢtur. Hedef doku yağ doku olduğu için yağ doku karĢılaĢtırma dokusu olarak belirlenmiĢtir. Daha sonra her doku için ayrı ayrı ΔCt değerleri bulunmuĢtur. Chrdl1 Ct değerinden Cyclophilin Ct değeri çıkarılmıĢtır. ΔΔCt değeri aĢağıdaki formülasyona göre hesaplanmıĢtır. ΔΔCt= ΔCtdoku – ΔCtyağ Daha sonra her doku için ayrı ayrı 2∆∆Ct değerleri hesaplanmıĢtır. Bulunan değer söz konusu dokulardaki gen ekspresyonunun yağ dokudan ne kadar farklı olduğunu göstermektedir. 3.2.9. Ġstatistik analizler Gerek Real Time PCR verileri gerekse insan ve fare ile ilgili mikroarray veri setlerinde bulunan farklı dokulardaki gen ekspresyon değerlerinin karĢılaĢtırmalı olarak değerlendirilmesinde tek yönlü varyans analizi (one-way ANOVA) kullanılmıĢtır. Tüm istatistik analizlerde SAS programının PROC GLM prosedürü kullanılmıĢtır (Anonymous, 2013d). Tez çalıĢmasında istatistik analizlerde kullanılan matematik model aĢağıda gösterilmektedir. Yij = μ + αi + εij Yij: i. dokuya ait j. gen ekspresyon değeri μ: Dokulara ait gen ekspresyon ortalaması, αi: i. dokunun etki miktarı, εij:Hata etkisini ifade etmektedir. 33 4. ARAġTIRMA SONUÇLARI VE TARTIġMA Günümüzde, mikroarray teknolojisi bilimsel araĢtırmalarda yoğun bir Ģekilde kullanılmaktadır. Bu teknoloji binlerce genin farklı biyolojik koĢullarda eĢ zamanlı olarak test edilmesine olanak sağlamaktadır. Bununla birlikte, burada zorlayıcı nokta binlerce gen arasından hedef genlerin nasıl seçilmesi gerektiğidir. Bu çalıĢma mikroarray ve GEO profil bilgilerinin etkin bir Ģekilde kullanılmasıyla, gerçek laboratuvar deneyleri yapılmadan önce yeni doku spesifik genlerin tespit edilmesine olanak sağlayan yeni bir metot sunmaktadır. Bu amaçla, insan ve farelerle ilgili ham mikroarray verileri GEO veri tabanından indirilmiĢtir (Anonymous, 2013a; Anonymous, 2013b). Aday genlerin belirlenmesi için adipoz doku hedef doku olarak belirlenmiĢtir. Daha sonra insan ve fare mikroarrayleri adipoz doku baz alınarak karĢılaĢtırmalı olarak analiz edilmiĢtir. Analiz sonuçları Chordin-like 1 (Chrdl1) geninin adipoz dokuyla ilgili aday bir gen olabileceğini göstermektedir. Bu sonucun bilimsel olarak desteklenmesi için insan ve fare mikroarray verilerinde Chrdl1 geni ile birlikte daha önce literatürde gösterilmiĢ adipoz spesifik genlerden FABP4 ve ATGL genleri ile ilgili istatistik analizler yapılmıĢtır. Sonuçlar Çizelge 4.1 ve 4.2’de gösterilmektedir. 34 Çizelge 4.1. Chrdl1, FABP4 ve ATGL genlerinin fare mikroarrayinde farklı dokulardaki gen ekspresyonu GEN ADĠPOZ KAS KALP AKCĠĞER KARACĠĞER BÖBREK DALAK P DEĞERĠ FABP4 2924.9a ± 443.944 198.6b ± 5.956 296.9b ± 34.924 86.1b ± 7.790 82.3b ± 0.988 118.0b ± 16.043 74.7b ± 5.626 P<0.0001 ATGL 4526.4a ± 388.379 785.7bc ± 82.611 1165.2b ± 78.833 453.9bc ± 36.512 327.2c ± 37.410 410.2c ± 34.468 195.8c ± 8.908 P<0.0001 CHRDL1 825.27a ± 129.857 99.81b ± 6.960 76.72b ± 2.002 110.23b ± 6.825 64.90b ± 1.262 97.79b ± 12.003 92.89b ± 3.453 P<0.0001 GSM252093 GSM252094 GSM252095 GSM252070 GSM252071 GSM252072 GSM252073 GSM252113 GSM252114 GSM252115 GSM252080 GSM252081 GSM252081 GSM252074 GSM252075 GSM252076 GSM252083 GSM252084 GSM252085 GSM252067 GSM252068 GSM252069 GSM NO GSM NO: adipoz, kas, kalp, akciğer, karaciğer, böbrek ve dalak dokularının biyolojik tekerrürlerini göstermektedir. Biyolojik tekerrürler adipoz, kalp, akciğer, karaciğer, böbrek ve dalak dokuları için (n=3) ve kas dokusu için (n=4) olarak gösterilmektedir. Sonuçlar ortalama ± standart hata olarak gösterilmektedir. Aynı satırda farklı harflerle gösterilen ortalamalar arasındaki farklar önemlidir (P<0.05). 35 Çizelge 4.2. Chrdl1, FABP4 ve ATGL genlerinin insan mikroarrayinde farklı dokulardaki gen ekspresyonu GEN FABP4 ATGL CHRDL1 ADĠPOZĠT KAS KALP AKCĠĞER KARACĠĞER BÖBREK 13.364.400a ± 503.100 2.838.150a ± 111.150 1.702.350a ± 86.950 446.400bc ± 130.800 367.800c ± 92.800 303.450b ± 191.450 317.400bc ± 153.700 853.150b ± 260.250 130.900b ± 80.600 1.099.850b ± 275.350 274.250c ± 54.750 315.400b ± 41.500 123.700c ± 74.000 299.600c ± 105.500 58.550b ± 27.050 189.500bc ± 58.100 357.850c ± 76.950 240.550b ± 24.550 GSM18975 GSM19013 GSM18951 GSM18949 GSM18953 GSM18955 GSM18976 GSM19014 GSM18952 GSM18950 GSM18954 GSM18956 P DEĞERĠ P<0.0001 P<0.0001 P<0.0001 GSM NO GSM NO: adipozit, kas, kalp, akciğer, karaciğer, böbrek ve dokularının biyolojik tekerrürlerini göstermektedir. Biyolojik tekerrürler adipozit, kas, kalp, akciğer, karaciğer, böbrek (n=2) olarak gösterilmektedir. Sonuçlar ortalama ± standart hata olarak gösterilmektedir. Aynı satırda farklı harflerle gösterilen ortalamalar arasındaki farklar önemlidir (P<0.05). 36 4.1. Bilimsel hipotezin kurulması Çizelge 4.1 ve Çizelge 4.2’ in incelenmesinden de anlaĢılabileceği gibi Chrdl1 geni insanlarda adipozitlerde ve farelerde adipoz dokuda iyi bilinen adipoz spesifik genlere (FABP4 ve ATGL) benzer Ģekilde diğer dokulara göre adipoz dokuda önemli derecede yüksek ekspresyon göstermektedir (P<0.0001). Gerek mikroarray verilerinin karĢılaĢtırmalı analizi gerekse yapılan istatistik analizler Chrdl1 geninin adipoz spesifik bir gen olma olasılığını güçlendirmektedir. Chrdl1 geninin türler arasındaki benzerliği (homolojisi) gen bankasında bulunan BLAST (Basic Local Alignment Search Tool) programıyla araĢtırılmıĢtır. Chrdl1 geninin sığırlardaki sekansı farklı canlı türleriyle karĢılaĢtırılmıĢtır. KarĢılaĢtırma sonuçlarına göre sığır Chrdl1 geni insan, fare ve ratlarda sırasıyla %87, %88 ve %87.5 benzerlik (homoloji) göstermektedir (Anonymous, 2013c). Bu durumda Chrdl1 geninin farklı canlı türlerinde aynı özelliği göstermesi söz konusu genin türler arasında korunmuĢ bir gen olduğuna iĢaret etmektedir. Elde edilen sonuçlar gen bankasında bulunan adipoz dokuyla ilgili farklı GEO profillerden elde edilen bilgilerle desteklenmiĢtir (Anonymous, 2013a; Anonymous, 2013b). ÇalıĢmada yararlanılan GEO profilller Çizelge 4.3’de gösterilmektedir. Tüm sonuçlar ve literatür bildiriĢleri değerlendirildiğinde Chrdl1 geninin adipoz spesifik bir gen olduğuyla ilgili bilimsel bir hipotez kurulmuĢtur. Kurulan hipotez, Dünya’da mikroarray sonuçlarının doğrulanmasında kullanılan en güvenilir moleküler yöntemlerin baĢında gelen Real Time PCR ile test edilmiĢtir. Tez çalıĢmasında kullanılan metot ġekil 4.1’de Ģematize edilmiĢtir. 37 İnsan Mikroarray Verisi Fare Mikroarray Verisi GEO veritabanından ham mikroarray verilerinin indirilmesi Aday genlerin mikroarray verilerinin karşılaştırmalı analiziyle belirlenmesi Aday genlerin seçim aşaması Hipotezin kurulması Hipotezin GEO profillleri ve literatür ile desteklenmesi Hipotezin laboratuvar denemeleriyle test edilmesi Sonuç ġekil 4.1. Tez çalıĢmasında izlenilen metot 38 Çizelge 4.3. Adipoz dokuyla ilgili GEO profiller GEO PROFĠL AÇIKLAMA GDS2818 Adipocytes vs. preadipocytes from subcutaneous and visceral fat depots (Mouse) GDS2659 Analysis of differentiating 3T3-L1 preadipocytes (Mouse) GDS734 Thiazolidinedione treatment of 3T3-L1 adipocytes (Mouse) GDS3688 Obese children: omental adipose tissue (Homo sapiens) GDS2366 Preadipocytes from anatomically separate fat depots (HG-U133A) (Homo sapiens) GDS1298 Fibroblast adipogenesis induced by EBF-1 and PPARgamma2 overexpression (Mouse) GDS2813 Comparison of brown and white adipose tissues (Mouse) GDS3102 Analysis of hearts and white adipose tissues of 28 month old male animals fed a calorie restricted diet since 4 months of age (Rattus norvegicus) GDS2319 Analysis of inguinal adipose tissue from genetically identical inbred C57BL/6J males exhibiting high or low weight gain after 4 weeks on a high saturated fat diet (Mouse) 4.2. Hipotezin Real Time PCR ile Test Edilmesi Bu çalıĢmada kullanılan metot, gerçek laboratuvar deneyleri yapılmadan önce doku spesifik aday genlerin belirlenmesine yönelik bilimsel hipotez kurulmasına olanak sağlamaktadır. Metodun son aĢamasını kurulan bilimsel hipotezin gerçek laboratuvar deneyleriyle test edilmesi oluĢturmaktadır. Bu amaçla kurulan bilimsel hipotez, Dünya’da mikroarray sonuçlarının doğrulanmasında kullanılan güvenilir ve tekrar edilebilirliği en yüksek moleküler yöntemlerin baĢında gelen Real Time PCR ile test edilmiĢtir. Farklı sığır dokularında Chrdl1 geniyle ilgili Real Time PCR’da yapılan gen ekspresyon denemeleri sonucunda oluĢturulan grafik ġekil 4.2’de gösterilmektedir. 39 ġekil 4.2. Chrdl1 geninin farklı sığır dokularındaki gen ekspresyonu Total RNA sığır adipoz, kalp, böbrek, akciğer, karaciğer, kas ve dalak dokularından izole edilmiĢtir. Chrdl1 geninin farklı sığır dokularındaki gen ekspresyon değerleri Real Time PCR’da ölçülmüĢtür (n=5). Grafikte görülen barlar ortalama ± standart hatayı temsil etmektedir. (*) iĢareti tek yönlü varyans analizini takiben yapılan Duncan önemlilik testinin sonuçlarına göre Chrdl1 geninin adipoz dokudaki ekspresyonunun diğer dokulara göre istatistik olarak önemli olduğunu göstermektedir (P<0.05). 40 ġekil 4.2’de gösterildiği üzere Chrdl1 geni adipoz dokuda diğer dokulara göre önemli derecede yüksek ekspresyon göstermektedir (P<0.05). Chrdl1 geni kalp, akciğer ve kas dokularında diğer dokulara göre biraz daha fazla ekspresyon göstermektedir. Bununla birlikte Chrdl1 geninin böbrek, karaciğer ve dalak dokularındaki ekspresyonu yok denecek kadar azdır. Elde edilen Real Time PCR sonuçları çalıĢma bünyesinde uygulanan metoda göre oluĢturulan bilimsel hipotezi desteklemektedir. Daha önce yapılan bilimsel çalıĢmalar Chrdl1 geninin geniĢ bir yelpazede farklı dokularda ekspresyon gösterdiğini ifade etmektedir (Coffnier ve ark., 2001; Nakayama ve ark., 2001; Sakuta ve ark.,. 2001; Bowen ve ark., 2009; Zheng ve ark., 2010). Literatür ve gen bankası tarandığında Chrdl1 geninin adipoz dokuda spesifik olduğuna dair hiç bir bulguya ve yayına rastlanılmamıĢtır. Tümüyle değerlendirildiğinde bu çalıĢma, Chrdl1 geninin adipoz dokuda spesifik bir gen olduğunu ortaya koyan ilk çalıĢmadır. Bu çalıĢmada uygulanan metodla, Chrdl1 geniyle birlikte adipoz dokuda spesifik yeni bir gen seti tespit edilmiĢtir. Bu genler arasında FABP4 (Yağ Asidi Bağlayıcı Protein 4) ve ATGL (Adipoz Trigliserit Lipaz) gibi oldukça iyi bilinen adipoz spesifik genler bulunmaktadır. Yapılan analizlere gore FABP4 ve ATGL genleri farelerde adipoz dokuda diğer dokulara göre oldukça yüksek gen ekspresyonu göstermektedir (Çizelge 4.1) (P<0.0001). Benzer Ģekilde FABP4 ve ATGL genleri insanlarda adipozitlerde diğer dokulara göre oldukça yüksek gen ekspresyonu göstermektedir (Çizelge 4.2) (P<0.0001). Bu çalıĢmada kullanılan metodun daha iyi anlaĢılabilmesi için FABP4 ve ATGL ve Chrdl1 geniyle ilgili yapılan çalıĢmalar örneklerle açıklanacaktır. 4.3. Yağ Asidi Bağlayıcı Protein 4 (FABP4) Gen bankasında FABP4 geni ile ilgili arama yapıldığında söz konusu gen ile ilgili GEO veritabanında 4.906 veri bulunduğu görülmektedir (Anonymous, 2013a; Anonymous, 2013b). Bu çalıĢmada, kullanılan metodla adipoz spesifik genlerin belirlenmesi amaçlandığı için aramanın daha da derinleĢtirilmesi gerekmektedir. Bu amaçla adipoz dokuyla ilgili “adipozit” ve “adipoz doku” anahtar kelimeleri gen bankasında bulunan GEO veritabanında taranmıĢtır. Taramalar sonucunda adipozit ve adipoz doku ile igili sırasıyla 2.786 ve 14.206 mikroarray verisi olduğu tespit edilmiĢtir. Mikroarray verileriyle bağlantılı GEO profillerinden elde edilen bilgiler bu çalıĢmada kullanılan metodun uygulanmasında önemli aĢamalardan biridir. Bu nedenle üzerinde 41 durulan genlerle ve hedef dokuyla ilgili ipuçlarını verecek GEO profillerin belirlenmesi büyük önem taĢımaktadır. Bu amaçla Çizelge 4.3’deki adipoz dokuyla ilgili bazı GEO profiller FABP4 geninin adipoz spesifik bir gen olduğunu açıklamak için kullanılacaktır. Buna ilaveten literatürden elde edilen bilgilerle kullanılan GEO profiller desteklenecektir. GDS2659, FABP4 gen ekspresyonunun adipozit hücrelerinin farklılaĢması sırasında artıĢ gösterdiğini ifade etmektedir (Cheung ve ark., 2007). Li ve ark. (2009), FABP4 geninin 3T3-L1 adipozit hücrelerinin farklılaĢması sırasında yüksek derecede ekspresyon gösterdiğini bildirmiĢtir. Bu bulgu aynı zamanda farklı araĢtırmacıların yaptıkları çalıĢmalarla teyit edilmektedir (Spiegelman ve Green 1983; Hunt ve ark., 1986; Samulin ve ark., 2008). GDS2818, FABP4 geninin adipozit hücrelerinde preadipozit hücrelerine (adipozit öncesi) göre daha fazla ekspresyon gösterdiğini ifade etmektedir (Gesta ve ark., 2006). GDS2818’i destekleyici Ģekilde Shin ve ark. (2008) ve Lee ve ark. (2009), yaptıkları çalıĢmalarda FABP4 geninin adipozit hücrelerinde preadipozit hücrelerine göre daha fazla ekspresyon gösterdiğini bildirmiĢlerdir. GDS3688, obez çocuklarda FABP4 geninin yüksek ekspresyon gösterdiğini ifade etmektedir (Aguilera ve ark., 2007). Ma ve ark. (2009), FABP4 ve PPAR-γ genlerinin obezite gösteren gruplarda obeziteye karĢı direnç gösteren gruplara göre yüksek derecede ekspresyon gösterdiğini bildirmiĢlerdir. GDS734, adipoz dokuda görev yapan transkripsiyonel faktörlerden PPAR-γ’nın FABP4 geninin adipozit hücrelerindeki ekspresyonunu tetiklediğini göstermektedir (Hsiao ve ark., 2004). Shin ve ark. (2009), PPAR-γ’nın FABP4 geninin promotoruna bağlanarak transkripsiyonunu tetiklediğini bildirmiĢtir. Petrovic ve ark. (2008), PPAR-γ reseptörüne bağlanan rosiglitazon ile muamele edilen kültürlerde FABP4 geninin muamale edilmeyen kontrol kültürlerine göre daha önce ortaya çıktığını göstermiĢtir. Genel olarak GEO profiller FABP4 geninin adipoz spesifik bir marker olduğunu göstermektedir. 42 4.4. Adipoz Trigliserit Lipaz (ATGL) Günümüzde, ATGL geni ile ilgili GEO veritabanında 4.588 veri bulunduğu görülmektedir (Anonymous, 2013a; Anonymous, 2013b). Bu veriler arasında ATGL geninin adipoz dokuda spesifik bir gen olduğunu gösteren GEO profillere bakıldığında; GDS2818, ATGL geninin adipozit hücrelerinde preadipozit hücrelerine göre daha fazla ekspresyon gösterdiğini ifade etmektedir (Gesta ve ark., 2006). Bu bulgu daha önce yapılmıĢ çalıĢmalarla doğrulanmaktadır. AraĢtırmacılar, ATGL geninin domuzlarda adipozit hücrelerinde, hücre kültürünün preadipozitlerce zengin kısmı olan stroma vaskular fraksiyona göre yüksek derecede ekspresyon gösterdiğini bildirmiĢlerdir. Ayrıca araĢtırmacılar, ATGL geninin tavuklarda bölünmüĢ adipoz hücrelerinde stroma vaskular fraksiyona kıyasla baskın bir Ģekilde yüksek ekspresyon gösterdiğini bildirmiĢlerdir (Deiuliis ve ark., 2008; Oh ve ark., 2011; Serr ve ark., 2011). GDS2366, ATGL gen ekspresyonunun adipoz hücrelerinin farklılaĢması sırasında artıĢ göstermektedir (Tchkonia ve ark., 2007). Kim ve ark. (2006), ATGL gen ekspresyonunun 3T3-L1 adipozit hücrelerinin farklılaĢması sırasında çarpıcı bir Ģekilde artıĢ gösterdiğini bildirmektedir. Deiuliis ve ark. (2008), ATGL gen ekspresyonunun domuz adipozit hücrelerinin farklılaĢması sırasında artıĢ gösterdiğini bildirmektedir. GDS3688, obez çocukların düĢük ATGL gen ekspresyonuna sahip olduklarını göstermektedir (Aguilera ve ark., 2007). Steinberg ve ark. (2007), obez örneklerin ATGL mRNA ekspresyonunu önemli derecede azalttığını göstermektedir (P<0.05). Jocken ve ark. (2007), insüline dirençli örneklerin ATGL mRNA ekspresyonunu insüline duyarlı örneklere nazaran azalttığını bildirmektedir. GDS1298, PPAR-γ (2)’nin fazla ekspresyon göstermesi durumunda NIH-3T3 embriyonik fibroblastların adipozitlere dönüĢmesi sırasında ATGL gen ekspresyonunu aktive ettiğini göstermektedir (Akerblad ve ark., 2005). Kershaw ve ark. (2007), yaptıkları araĢtırmalarda PPAR-γ’nın canlı ve laboratuvar ortamlarında ATGL mRNA ekspresyonunu düzenlediğini bildirmiĢtir. Sonuç olarak GEO profilleri, ATGL geninin adipoz spesifik olmasına ve ATGL gen ekspresyonunun adipoz doku geliĢimi üzerine etkilerine dikkat çekmektedir. 43 4.4. Chordin-like 1 (Chrdl1) GEO veri tabanına bakıldığında Chrdl1 ile ilgili 3.504 veri bulunduğu görülmektedir (Anonymous, 2013a; Anonymous, 2013b). Bu veriler değerlendirildiğinde Chrdl1 ve adipoz dokuyla ilgili sadece 31 GEO profil olduğu görülmektedir. Bu GEO profillerden elde edilen bilgiler süzüldüğünde sayı daha da azalmaktadır. Chrdl1 ve adipoz dokuyla ilgili GEO profiller değerlendirildiğinde; GDS2813, Chrdl1 geninin beyaz adipoz dokuda kahverengi adipoz dokuya göre ekspresyonunun daha fazla olduğunu göstermektedir (Seale ve ark., 2007). GDS3102, diyetlerde kalorik kısıtlamanın Chrdl1 gen ekspresyonunu azalttığını göstermektedir (Linford ve ark., 2007). GDS2366, Chrdl1 gen ekspresyonunun bölünüp olgun adipozitlere dönüĢen preadipozitlerde bölünmeyen preadipozitlere göre çok daha fazla olduğunu göstermektedir (Tchkonia ve ark., 2007). GDS2319, yüksek ağırlık kazancı gösteren bireylerin düĢük ağırlık kazancı gösteren bireylere göre yüksek Chrdl1 gen ekspresyonuna sahip olduklarını göstermektedir (Koza ve ark., 2006). Chrdl1 geni ile ilgili çalıĢmanın kaynak araĢtırması bölümünde belirtilen literatürler ve GEO profiller değerlendirildiğinde Chrdl1 geni ve adipoz dokuyla ilgili bazı ipuçlarına ulaĢılmaktadır. Bunun ötesinde daha öncede ifade edildiği gibi yapılan istatistik analizler gerek insan gerekse fare mikroarray verilerinde Chrdl1 geninin adipoz dokuda yüksek derecede ekspresyon gösterdiğini ortaya koymaktadır (P<0.001). Chrdl1 geninin farklı dokularda ekspresyon gösterdiği bildirilmesine rağmen adipoz dokudaki ekspresyonuyla ilgili herhangi bir bildiriĢ bulunmamaktadır. Bu çalıĢmada kullanılan metot sayesinde Chrdl1 geninin adipoz dokuya spesifik bir gen olduğuyla ilgili kurulan hipotez baĢarıyla test edilmiĢtir. Bunun ötesinde kullanılan metot kapsamında yapılan karĢılaĢtırmalı mikroarray analizleri Chrdl1 geninin adipoz dokuda Ģu ana kadar bildirilen bir çok adipoz spesifik genden (adipokin) daha fazla ekspresyon gösterdiğini ortaya koymaktadır. Bu durumda Chrdl1 geninin, diğer adipoz spesifik genler gibi adipoz dokuda önemli bir fonksiyon gösterme olasılığı oldukça yüksek olmaktadır. 44 5. SONUÇLAR VE ÖNERĠLER 5.1. Sonuçlar Bu çalıĢma, gen bankasında bulunan mikroarray verilerinin karĢılaĢtırmalı olarak detaylı analizi yapılmak suretiyle doku spesifik aday genlerin ortaya çıkarılmasına olanak sağlayan yeni bir metot sunmaktadır. Bu metot laboratuvar deneyleriyle baĢarılı bir Ģekilde test edilerek uygulanabilirliği ispat edilmiĢtir. Bu metotun kullanılması sayesinde literatüre katkı sağlıyacak yeni bir dizi adipoz spesifik aday gen belirlenmiĢtir. Adipoz dokunun sahip olduğu biyolojik önem dikate alındığında araĢtırma sonuçlarının önemli hedeflere ulaĢması beklenmektedir. Bilim adamları gerek hayvanlarda gerekse insanlarda yaptıkları çalıĢmalarla adipoz dokunun vücutta nasıl fonksiyon gösterdiğini ve burada ne tür moleküler mekanizmaların devreye girdiğini ortaya çıkarmayı hedeflemektedir. Bu nedenle bu çalıĢmada belirlenen adipoz spesifik gen ve diğer aday genlerle ilgili ileride yapılacak olan çalıĢmalar adipoz doku geliĢiminin moleküler mekanizmasının aydınlatılmasında, dolayısıyla adipoz dokunun vücutta gösterdiği fonksiyonların daha iyi anlaĢılması bakımından büyük önem taĢımaktadır. Son yıllarda yapılan çalıĢmalar neticesinde, adipoz dokuda meydana gelen anormalliklerin insanlarda obezite, tip 2 diyabet, kalp hastalıkları ve bazı kanser türleriyle iliĢkisi olduğu düĢünüldüğünde bu çalıĢmada tespit edilen adipoz spesifik gen ve diğer aday genlerle ilgili ileride yapılacak olan çalıĢmaların söz konusu hastalıkların tedavisine yardımcı olarak insan sağlığına önemli katkılar sağlaması kuvvetle muhtemeldir. Çiftlik hayvanlarında aĢırı yağlanma gerek önemli sağlık problemlerine gerekse ekonomik bakımdan önemli kayıplara neden olmaktadır. Bu çalıĢmayı izleyen araĢtırmalarda tespit edilen genlerin markere dayalı seleksiyon kriteri olarak kullanılıp kullanılamayacağı ile ilgili yapılacak olan denemelerin olumlu sonuç vermesi durumunda çiftlik hayvanlarında üretim kalitesinin arttırılması ve daha sağlıklı ve ekonomik üretim yapılması konularında ciddi mesafeler kaydedilebileceği düĢünülmektedir. Bu çalıĢma, doku spesifik aday genlerin belirli bir strateji takip edilerek belirlenmesine yönelik yeni bir metot sunmaktadır. Bu metotun sunduğu bazı önemli avantajlar bulunmaktadır. Ġlk olarak bu metot gerçek laboratuvar deneyleri 45 yapılmaksızın aday genlerin tespitine olanak sağlamaktadır. Bu durum ciddi bir zaman kaybını önlemektedir. Öne çıkan diğer bir avantaj ise bu metodun kullanılması durumunda gereksiz yere yapılan araĢtırma harcamalarının önüne geçilmesidir. Öyleki bilim adamlarının bu metodu izlemesi durumunda halihazırda gen bankasında daha önce hazırlanmıĢ mikroarray verilerini kullanmaları sağlanarak bu çalıĢmaları gereksiz yere tekrar edip para ve zaman kaybına uğramalarının önüne geçilebilecektir. Bu çalıĢmanın sunduğu bir önemli avantaj ise gen bankasının etkili olarak kullanımı, verilerin süzülmesi ve analizi ve sonuçların değerlendirilmesi konularında sağladığı öğretici bilgilerdir. Bu çalıĢmada, adipoz doku insanlar ve hayvanlar için sahip olduğu biyolojik önemden dolayı hedef doku olarak metodun uygulanmasında kullanılmıĢtır. Bununla birlikte bilim adamlarının aynı metodu farklı dokularda denemesi durumunda söz konusu dokulara spesifik yeni genleri tespit edebilmesi kuvvetle muhtemeldir. Bu metodun farklı dokularda denenmesi durumunda elde edilecek sonuçların literatüre önemli katkılar sağlaması beklenmektedir. 5.2. Öneriler Dünyada adipoz dokuyla ilgili yapılan bilimsel çalıĢmalara bakıldığında, araĢtırmaların insan ve hayvanlarda sağlık, çiftlik hayvanlarında önemli verim özelliklerinin geliĢtirilmesi ve bu özelliklerin moleküler mekanizmalarının ortaya çıkarılmasına yönelik markere dayalı seleksiyon yöntemlerinin kullanılması konularında yoğunlaĢtığı görülmektedir. Tez çalıĢması süresince yapılan araĢtırmalar, adipoz dokuyla ilgili yapılan bilimsel çalıĢmaların kapsamı ve ülke koĢulları değerlendirildiğinde öneriler kısmının, ulusal karkas derecelendirme sisteminin kurulması, markere dayalı seleksiyon yöntemlerinin kullanılması ve sağlıklı beslenme konusunda toplumsal bilincin arttırılması olarak 3 ana baĢlık altında toplanmasında fayda görülmektedir. 5.2.1. Ulusal karkas derecelendirme sisteminin kurulması Dünyada karkas özelliklerinin sınıflandırılması ve bu sınıflandırmaya göre fiyatlandırılmasına yönelik ülkelerin uyguladıkları karkas derecelendirme sistemleri incelendiğinde, uygulamada az da olsa farklılıklar olmasına rağmen karkas 46 derecelendirme sistemlerinin et verim derecesi (yield grade) ve kalite derecesi (quality grade) olmak üzere iki önemli unsurdan oluĢtuğu görülmektedir. Et verim derecesi karkastaki yağlanmaya bağlı olarak belirlenirken, kalite verim derecesi de ette görülen mozaikleĢmeye bağlı olarak, etin gevreklik, sululuk ve tad özelliklerine göre belirlenmektedir. Et verim derecesi ve kalite derecesinin birlikte değerlendirilmesi sonucunda oluĢturulan skalaya göre ette fiyatlandırma yapılıp pazara sunulmaktadır (ZoBell ve ark., 2005). BaĢta ABD ve Avrupa Birliği ülkelerinde karkas derecelendirme sisteminin kurulumu, yönetimi ve denetimini tarım bakanlığı üstlenmektedir. Türkiye de malesef sığır karkaslarının sınıflandırılıp fiyatlandırılmasına yönelik bir karkas derecelendirme sistemi bulunmamaktadır. Bu konuyla ilgili günümüze kadar yapılmıĢ çalıĢmalara bakıldığında, Türk Standart Enstitüsünün 15.03.1988 tarihinde yürürlüğe giren TSE 383 nolu kasaplık sığır baĢlıklı standardı tamamen subjektif kriterlere göre yapılan bir derecelendirme olarak ortaya konulmuĢtur. Öyleki söz konusu standarda göre karkastaki etlilikle ilgili hususlar karkasın uzunluk ve derinliğine, butların yuvarlak, uzun ve dolgun oluĢuna, omuzların kalın ve belirgin oluĢuna ve boyun ve bacakların kısalık ve dolgunluğuna göre tam etli, etli ve az etli olarak üç grupta toplanmaktadır. Karkasta yağlılık ise kabuk yağının yayılıĢ alanı, kalınlığı ve rengi, iç yağların yani göğüs, karın ve leğen yağlarının kalınlığı ve salkım görüntüsü dikkate alınarak tam yağlı, yağlı ve az yağlı olmak üzere üç grupta toplanmakta olup ancak gruplara yönelik herhangi bir fiyatlandırma yapılmamıĢtır. (Anonim, 2011b) TSE 383 nolu standardın yürürlükten kalkmasından 2007 yılına kadar Türk Standart Enstitüsünün "Sığır Gövde Etleri (Karkas) Standardı (TS 668)” uygulanmıĢ söz konusu standartta 21.06.2007 yılında yürürlükten kaldırılmıĢtır. Tarım ve KöyiĢleri Bakanlığınca yayınlanan "Kasaplık Gövde Etlerinde Standart Uygulaması; Sığır Gövde Etleri (Karkas) Standardını (TS 668)” yeniden düzenleyerek 01.01.2010 tarihinden itibaren yapılan kesimlerde böbrek yağları, pelvis boĢluğu yağları, salkım ve fıtık yağlarının sıcak tartıdan önce karkastan uzaklaĢtırmasını öngörmektedir (Anonim, 2010b, Anonim, 2011c). Karkas derecelendirme sisteminin getirdiği en büyük avantaj tüketiciyi aldığı ürünle ilgili bilgilendirerek kalitesiz ürüne fazla ücret ödemesini önlemesidir. Türkiye de bu Ģekilde bir uygulamanın olmamasından dolayı tüketici kaliteli ve kalitesiz ürüne eĢit ücret ödemektedir. Örneğin 1.5 yaĢlı tosundan elde edilen biftek veya bonfile birim fiyatı ile 4.5 yaĢlı boğadan veya sığırdan elde edilen aynıdır. Ücretlendirme herhangi bir sınıflandırma olmaksızın biftek veya bonfile diye yapılmaktadır. Karkas derecelendirme 47 sisteminin getirdiği bir diğer önemli avantaj üreticiyi kaliteli üretim yapmaya teĢvik etmesidir. Öyleki daha kaliteli üretim yapmak için çaba gösteren üretici karkas derecelendirme sistemi sayesinde ürününün pazarda daha yüksek fiyata alıcı bulmasıyla emeğinin karĢılığını almaktadır. Üreticiler söz konusu fiyat avantajından yararlanmak amacıyla üretimlerinde geleneksel üretimi bir yana bırakıp profesyonel ekiplerin nezaretinde daha modern ve bilimsel üretim yapma gayreti göstermektedir. Karkas derecelendirme sisteminde fiyatlandırma karkasın yağlılık durumuna göre değiĢmekte olup karkasta yağlanma arttıkça ters orantılı olarak karkasın fiyatlandırma değeri düĢmektedir. Bu durum tüm dikkatleri yağ dokunun üzerine çekerek bilimsel bir farkındalık oluĢturmaktadır. Öyleki karkastaki kabuk yağı, böbrek üstü salkım yağları ve iç organlar etrafındaki ekonomik değeri olmayan yağ miktarı azaltılırken karkasta ekonomik değeri olan mozaikleĢmeyi arttırıcı kas içi yağlar nasıl arttırılabilir sorusu bilim adamlarının ısrarla üzerinde durduğu bir konu haline gelmiĢtir. Genotip ve beslenme karkasta yağlanmayı arttıran iki önemli faktördür. Besi döneminde aynı yaĢta, aynı ırkta ve aynı canlı ağırlıktaki hayvanların besi performansı farklı olmaktadır. Aynı özelliklere sahip hayvanların aynı rasyonlarla beslenmesi durumunda bazı hayvanlarda karkasta aĢırı yağlanma görülürken bazılarının vücut kondüsyonu ideal seviyelerde olmaktadır. Burada öne çıkan özellik genotip olmaktadır. Yağ dokunun oluĢmasının genetik mekanizmasının ortaya çıkarılması için bilim adamları yağ dokunun geliĢmesini etkileyen genleri araĢtırmaktadır. Ulusal karkas derecelendirme sisteminin oluĢturulmasının yağ dokunun öneminin anlaĢılması adına gerek üretici ve tüketici, gerekse bilimsel camiada bir farkındalık oluĢturması beklenmektedir. Tümüyle değerlendirildiğinde, Gıda, Tarım ve Hayvancılık Bakanlığı’nın Türkiye’deki üretici ve tüketici taleplerini göz önünde bulundurarak ulusal karkas derecelendirme sisteminin oluĢturulmasına yönelik çalıĢmalara vakit kaybetmeden baĢlaması ülke menfaatleri açısından önem taĢımaktadır. 5.2.2. Markere dayalı seleksiyon yönteminin kullanılması Son yıllarda çiftlik hayvanlarında önemli verim özelliklerinin iyileĢtirilmesine yönelik markere dayalı seleksiyon yöntemlerinin kullanılması giderek yaygınlaĢmaktadır. Markere dayalı seleksiyon yönteminin kullanılması seleksiyonda isabet derecesini arttırarak generasyonlar arası süreyi kısaltmaktadır. Bu yöntemde baĢarı, üzerinde durulan özellikle ilgili doğru markerlerin tespit edilmesine bağlıdır. 48 Yağ dokunun geliĢmesi üzerinde etkili markerlerin belirlenmesi bu dokuyla ilgili moleküler gizemin çözülmesine katkıda bulunacaktır. Yağ doku ile kas doku arasında geliĢimsel olarak bir ters orantı bulunmaktadır. Daha açık bir ifadeyle karkasta yağlılık arttıkça kaslılık o derecede azalmaktadır. Daha öncede ifade edildiği üzere kırmızı ette kas içi yada kas arasında görülen yağlanma karkasın ekonomik değerini arttırırken kabuk yağı, böbrek ve iç organların etrafındaki yağlar karkasın ekonomik değerini düĢürmekle kalmayıp kesim sonrasında karkastan ayrılıp atılmalarından dolayı karkas randımanını düĢürmektedir. Bu durumda karkasta yağlanmanın istenilen bölgelerde arttırılması ekonomik açıdan önem taĢımaktadır. Karkasta mozaikleĢmeyi arttırıcı genetik markerlerin belirlenmesine yönelik çalıĢmalar artarak devam etmektedir. Bu çalıĢmaların önemli bir çoğunluğu FABP4 geninin çiftlik hayvanlarında mozaikleĢme özelliğiyle ilgili genetik bir marker olarak kullanılabileceğine iĢaret etmektedir. Bununla birlikte bugüne kadar yağ dokunun genetik yapısı üzerine FABP4 geni gibi majör etkili yaklaĢık olarak sadece 20 gen tespit edilebilmiĢtir. Yağ doku ile ilgili yapılan araĢtırmalarda markere dayalı seleksiyon yöntemlerinin kapsamının geniĢ tutulması, çok daha fazla markerin tespit edilmesine dolayısıyla yağ dokunun genetik yapısıyla ilgili daha fazla bilgiye ulaĢılmasını sağlayacaktır. 5.2.3. Sağlıklı beslenme konusunda toplumsal bilincin arttırılması Son yıllarda kırmızı et tüketiminin insan sağlığı üzerinde olumsuz etkileri olduğu yönünde çıkan medya haberleri ve bilimsel çevrelerden yapılan bazı açıklamalar toplumda kırmızı et tüketimine karĢı ciddi bir ön yargı oluĢturmuĢtur. Kırmızı et tüketimiyle ilgili bu ön yargıyı kaldırmak için tüketicilerin kırmızı et tüketimiyle ilgili doğru bilgilendirilmesi gerekmektedir. Kırmızı et vücudun ihtiyacı olan esansiyel amino asitler, demir, çinko, B12 vitamini ve vücutta kolayca emilimi sağlanan fosfor ve kalsiyumca zengin bir hayvansal besindir (Anonim, 2012b). Esansiyel amino asitler vücüt tarafından sentezlenemeyen ve mutlaka dıĢardan diyetle alınması gereken amino asitler olarak tanımlanmaktadır (Türkmen, 2003). Esansiyel amino asitler arasında arginin, histidin, izolösin, lösin, lizin, metiyonin, fenilalanin, treonin, triptofan ve valin amino asitleri bulunmaktadır. Hayvansal protein kaynağı olarak kırmızı et bütün bu esansiyel amino asitleri içermektedir (Anonim, 2013d). 49 YetiĢkin bir insanın ihtiyacı olan esansiyel amino asit miktarları Çizelge 5.1’de gösterilmektedir (Kaya, 1996). Çizelge 5.1. YetiĢkin bir insanın ihtiyacı olan esansiyel amino asit miktarları (Kaya, 1996) Esansiyel amino asidi Lysine Tryptophan Phenylalanine Threonine Valine Methionine Leucine Isolecine Miktar (g/gün) 0.8 1.4 1.1 0.5 0.8 1.1 1.1 0.7 Normal koĢullarda sağlıklı beslenen bir insanın her 1 kg canlı ağırlığına karĢılık 1g protein tüketmesi gerektiği ifade edilmektedir. Tüketilmesi gereken 1g protein’in % 42-50’lik kısmının hayvansal proteinden karĢılanması önem arz etmektedir. Kırmızı etin esansiyel aminoasit içeriği ve yetiĢkin bir insanın günlük ihtiyaçları göz önünde bulundurulduğunda günlük 150 g kırmızı et tüketimiyle günlük hayvansal protein ihtiyacının karĢılanabileceği bildirilmektedir (Kaya, 1996). Ne yazıkki kırmızı et son yıllarda biyolojik öneminden daha çok kolesterol, kalp hastalıkları, obezite, hipertansiyon gibi rahatsızlıklara neden olduğu ile ilgili yapılan haberlerle gündemde bulunmaktadır. Bu durum tüm dünya genelinde kırmızı et tüketiminde azalmalara yol açmaktadır. Burada dikkat çekilmesi gereken esas nokta dengeli beslenme olmalıdır. Vücut homeostazisinin korunması için bitkisel ve hayvansal proteinlerin yeterli miktarda alınması gerekmektedir. Tek yönlü beslenmenin önemli sağlık problemlerini getireceği göz ardı edilmemelidir. Öyleki çocuklarda sadece tahıla dayalı beslenmenin kwashiorkor olarak bilinen bir protein eksikliğine bağlı büyüme gecikmesi, apati, ödem, saç ağarması ve dökülmesi gibi sağlık sorunlarına neden olduğu belirtilmektedir (Anonim, 2013d). Üzerinde durulması gereken bir diğer konu ise kolesterol, kalp hastalıkları, obezite, hipertansiyon gibi rahatsızlıklara neden olan etmenlerdir. Söz konusu hastalıkların ortaya çıkmasında genetik yapı, dengesiz beslenme ve hareketsiz yaĢam gibi etkili bir çok faktör bulunmaktadır. Kırmızı et tüketimiyle ilgili esas öne çıkarılması gereken konu aĢırı yağlı kırmızı et tüketiminin sınırlandırılmasıdır. Az önce ifade edilen kolesterol, kalp hastalıkları, obezite, hipertansiyon gibi rahatsızlıkların 50 oluĢumunda diğer faktörlerle birlikte yağ dokunun önemli etkisi vardır. Sonuç itibariyle aĢırı yağlı kırmızı et tüketiminin söz konusu rahatsızlıklara neden olan faktörlerden biri olduğu; bununla birlikte kırmızı etin diyette yeterli alınması durumunda herhangi bir sağlık problemine neden olmadığı aksine vücut ihtiyaçlarının karĢılanması açısından önem taĢıdığıyla ilgili toplumsal bilincin arttırılmasında fayda görülmektedir. 51 KAYNAKLAR Anonim, 2010a, Üniversitemizde sıvı azot üretimi baĢladı, http://www.bolununsesi.com [Ziyaret Tarihi: 30 Haziran 2010]. Anonim, 2010b, Kasaplık Gövde Etlerinde Standart Uygulaması, http://www.stb.org.tr/Duyuru-Detay/?id=73 [Ziyaret Tarihi: 04 Nisan 2012]. Anonim, 2011a, Porselen Havan, http://www.deneycell.com [Ziyaret Tarihi: 20 Nisan 2011]. Anonim, 2011b, TS 383 kasaplık sığır, https://intweb.tse.org.tr [Ziyaret Tarihi: 15 Nisan 2011]. Anonim, 2011c, TS 668 Kasaplık sığır gövde etleri (karkas), https://intweb.tse.org.tr [Ziyaret Tarihi: 15 Nisan 2011]. Anonim, 2012a, ĠletiĢim, http://www.yilet.com/index-19.htm [Ziyaret Tarihi: 04 Nisan 2012]. Anonim, 2012b, Kalp Hastalarına kurbanda 'kırmızı et uyarısı, http://www.memurlar.net/haber/299161/ [Ziyaret Tarihi: 07 Nisan 2012]. Anonim, 2013a, Karkas Randımanını Etkileyen Faktörler,Yenilebilir Et Miktarını Etkileyen Faktörler, http://zooteknik.blogspot.com/2013/05/karkas-randmannetkileyen.html [Ziyaret Tarihi: 01 ġubat 2013]. Anonim, 2013b, Lipoatrofi (lipodistrofi) Nedir, http://www.birdunyabilgi.org/lipoatrofi-lipodistrofi-nedir [Ziyaret Tarihi: 10 Nisan 2013]. Anonim, 2013c, Yağ dokusu, http://tre.docdat.com/docs/5168/index-10535.html [Ziyaret Tarihi: 10 Nisan 2013]. Anonim, 2013d, Proteinler, http://www.msxlabs.org/forum/saglikli-yasam/145726proteinler-ve-beslenmedeki-onemi.html [Ziyaret Tarihi: 10 Nisan 2013]. Anonymous, 2010, Adipogenesis, http://en.wikipedia.org/wiki/Adipogenesis [Ziyaret Tarihi: 07 Temmuz 2010]. Anonymous, 2011a, Fatmouse, http://en.wikipedia.org/wiki/File:Fatmouse.jpg [Ziyaret Tarihi: 10 ġubat 2011]. Anonymous, 2011b, ND-1000 Spectrophotometer T009‐Technıcal Bulletin, https://bioscibatzerlab.biology.lsu.edu/Genomics/documentation/3130_NanoDrop _tips.pdf [Ziyaret Tarihi: 29 Mart 2011]. Anonymous, 2012a, Angus cattle, http://commons.wikimedia.org/wiki/File:Angus_cattle.jpg [Ziyaret Tarihi: 02 Mart 2012]. 52 Anonymous, 2012b, Tm Calculator, http://www6.appliedbiosystems.com/support/techtools/calc/ [Ziyaret Tarihi: 27 Nisan 2012]. Anonymous, 2012c, Vector NTI® Software, http://www.invitrogen.com/site/us/en/home/Products-and Services/Applications/Cloning/vector-nti-software.html [Ziyaret Tarihi: 27 Nisan 2012]. Anonymous, 2013a, GEO profiles, www.ncbi.nlm.nih.gov/geoprofiles/ [Ziyaret Tarihi: 12 Mayıs 2013]. Anonymous, 2013b, GEO DataSets, www.ncbi.nlm.nih.gov/geoprofiles/ [Ziyaret Tarihi: 12 Mayıs 2013]. Anonymous, 2013c, National Center for Biotechnology Information Database, http://www.ncbi.nlm.nih.gov [Ziyaret Tarihi: 3 Mart 2013]. Anonymous, 2013d, The GLM Procedure, http://support.sas.com [Ziyaret Tarihi: 15 Nisan 2013]. Aguilera, C. M, Tofe, I., Suarez, A., Gomez Llorente, C., Cañete, R. Gil, A., 2007, Differential gene expression in omental adipose tissue from obese children, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE9624 [Ziyaret Tarihi: 05 Haziran 2011]. Akerblad, P., Mansson, R., Lagergren, A., Westerlund, S., Basta, B., Lind, U., Thelin, A., Gisler, R., Liberg, D., Nelander, S., Bamberg, K. Sigvardsson, M., 2005, Gene expression analysis suggests that EBF-1 and PPARgamma2 induce adipogenesis of NIH-3T3 cells with similar efficiency and kinetics, Physiological Genomics, 23(2), 206-16. Avram, A. S., Avram, M. M. and James, W. D., 2005, Journal of the American Academy of Dermatology, 671-683. Bai, L, Pang, W. J, Yang, Y. J. and Yang, G. S., 2008, Modulation of Sirt1 by resveratrol and nicotinamide alters proliferation and differentiation of pig preadipocytes, Molecular and Cellular Biochemistry, 307(1-2), 129-40. Bjorbaek, C. and Kahn, B. B., 2004, Leptin signaling in the central nervous system and the periphery, Recent Progress in Hormone Research, 59, 305-31. Bowen, N. J., Walker, L. D., Matyunina, L. V., Logani, S., Totten, K. A., Benigno, B. B. McDonald, J. F., 2009, Gene expression profiling supports the hypothesis that human ovarian surface epithelia are multipotent and capable of serving as ovarian cancer initiating cells, BMC Medical Genomics, 29, 2, 71. 53 Buchanan, F.C., Van Kessel A. G., Waldner C., Christensen, D. A., Laarveld, B. Schmutz, S. M., 2003, Hot topic: an association between a leptin single nucleotide polymorphism and milk and protein yield, Journal of Dairy Science, 86(10), 3164-6. Castracane, V. D. and Henson, M. C., 2007, The Obese (ob/ob) Mouse and the Discovery of Leptin, Endocrine Updates, 25, 1-9. Chan, K.W., Lim, P. L., Tam, F. C. H., Li, E.T.S. Lim, B. L., 2000, Isolation of leptinbinding peptides from a random peptide phage library, Journal of Peptide Research, 55, 318-324. Chebel, R. C., Susca, F. Santos J. E., 2008, Leptin genotype is associated with lactation performance and health of Holstein cows, Journal of Dairy Science, 91(7), 2893900. Cheung, K. J, Tzameli, I., Pissios, P., Rovira, I., Gavrilova, O., Ohtsubo, T., Chen, Z., Finkel, T., Flier, J. S. Friedman, J. M., 2007, Xanthine oxidoreductase is a regulator of adipogenesis and PPARgamma activity, Cell Metab, 5(2), 115-28. Cho, S., Park, T. S., Yoon, D. H., Cheong, H. S., Namgoong, S., Park, B. L., Lee, H. W., Han, C. S., Kim, E. M., Cheong, I. C,, Kim, H. Shin, H. D., 2008, Identification of genetic polymorphisms in FABP3 and FABP4 and putative association with back fat thickness in Korean native cattle, BMB reports Online, 41(1), 29-34. Cinti, S., 2005, The adipose organ, Prostaglandins, Leukotrienes and Essential Fatty Acids, 73, 9-15. Clark, D. L., 2008, Transcription Profiles of Differentially Marbled Beef Cattle. Graduate College of the University of Illinois at Urbana Champaign, Master Thesis. Clempson, A. M., Pollott, G. E., Brickell, J. S., Bourne, N. E, Munce, N. Wathes D. C., 2011, Evidence that leptin genotype is associated with fertility, growth, and milk production in Holstein cows, Journal of Dairy Science, 94(7), 3618-28. Coffinier, C., Tran, U., Larrain, J. De Robertis, E. M., 2001, Neuralin-1 is a novel Chordin-related molecule expressed in the mouse neural plate, Mechanisms of Development, 100(1), 119-22. Cornelius, P., MacDougald, O. A. and Lane, M. D., 1994, Regulation of adipocyte development, Annual Review of Nutrition, 14: 99-129. Cui, H., Meng, Q., Qu, Y., Liu, H., Feng, C., Wang, H., Liu, Y., Zan, L. Li, N. 2011, Two novel missense mutations in bovine ATGL gene and their association with economic traits in Qinchuan cattle, African Journal of Biotechnology, 10, 23532359. 54 Da Silva, R. C., Ferraz, J. B., Meirelles, F. V., Eler, J. P., Balieiro, J. C., Cucco, D. C., Mattos, E. C., Rezende, F. M. Silva, S. L., 2012, Association of single nucleotide polymorphisms in the bovine leptin and leptin receptor genes with growth and ultrasound carcass traits in Nellore cattle, Genetics and Molecular Research, 11(4), 3721-8. Dai, L. H., Xiong, Y. Z., Jiang, S. W. Chen, J. F. 2011, Molecular characterization and association analysis of porcine adipose triglyceride lipase (PNPLA2) gene, Molecular Biology Reports, 38, 921-927. Deiuliis, J. A., Shin, J., Bae, D., Azain, M. J., Barb, R. Lee, K., 2008, Developmental, hormonal, and nutritional regulation of porcine adipose triglyceride lipase (ATGL), Lipids, 43, 215-25. DeVuyst, E. A., Bauer, M. L., Cheng, F. C., Mitchell, J. Larson, D., 2008, The impact of a leptin gene SNP on beef calf weaning weights, Animal Genetics, 39(3), 2846. Falcão Pires, I., Castro Chaves, P., Miranda Silva, D., Lourenço, A. P. Leite Moreira, A. F., 2012, Physiological, pathological and potential therapeutic roles of adipokines, Drug Discovery Today, 17(15-16), 880-9. Fernyhough, M. E., E. Okine, G. Hausman, J. L. and Vierck, M. V. Dodson., 2007, Invited review: PPAR-gamma and GLUT-4 expression as differentiation markers for preadipocyte conversion to become an adipocyte, Domestic Animal Endocrinology, 33, 367-378. Flier, J. S., 1998, What’s in a Name? In Search of Leptin’s Physiologic Role, Journal of Clinical Endocrinology and Metabolism, 83(5), 1407-1412. Friedman, J. M. and Halaas, J. L., 1998, Leptin and the regulation of body weight in mammals, Nature, 395, 764-770. Furuhashi, M. and Hotamisligil, G. S., 2008, Fatty acid-binding proteins: role in metabolic diseases and potential as drug targets, Nature Rev. Drug Discov, 7, 489503. Galic, S., Oakhill, J. L. and Steinberg, G. R., 2010, Adipose tissue as an endocrine organ, Molecular and Cellular Endocrinology, 316, 129-139. Gerbens, F., Jansen, A., Van Erp, A. J. M., Harders, F., Meuwissen, T. H. E., Rettenberger, G., Veerkamp, J. H. and Te Pas, M. F.W., 1998, The adipocyte fatty acid-binding protein locus: characterization and association with intramuscular fat content in pigs, Mammalian Genome, 9, 1022-1026. Gesta, S., Blüher, M., Yamamoto, Y., Norris, A. W., Berndt, J., Kralisch, S., Boucher, J., Lewis, C. Kahn, C. R., 2006, Evidence for a role of developmental genes in the origin of obesity and body fat distribution, Proceedings of the National Academy of Sciences, 103(17), 6676-81. 55 Grundy, S. M., Brewer, H. B., Cleeman, J. J. I., Smith, S. C. and Lenfant, C., 2004, Definition of Metabolic Syndrome, Arteriosclerosis Thrombosist Vascular Biology, 24, e13-e18. Haunerland, N. H. and Spener, F., 2004, Fatty acid-binding proteins insights from genetic manipulations, Progress in Lipid Research, 43, 328-349. Hoashi, S., Hinenoya. T., Tanaka, A., Ohsaki, H., Sasazaki, H., Taniguchi, M., Oyama, K., Mukai, M. and Mannen. H., 2008, Association between fatty acid compositions and genotypes of FABP4 and LXR-alpha in Japanese Black cattle, BMC Genetics, 9, 84. Holm, C., 2003, Molecular mechanisms regulating hormone-sensitive lipase and lipolysis, Biochemical Society Transactions, 31, 6. Hou, X, M., Li, Q, Z. and Huang, T, Y., 2010, Microarray analysis of gene expression profiles in the bovine mammary gland during lactation, Science China Life Sciences, 53(2), 248-256. Hsiao, A., Worrall, D. S., Olefsky, J. M. Subramaniam, S., 2004, Variance-modeled posterior inference of microarray data: detecting gene-expression changes in 3T3L1 adipocytes, Bioinformatics, 20(17), 3108-27. Hunt, C. R., Ro, J. H., Dobson, D. E., Min, H. Y. Spiegelman, B. M., 1986, Adipocyte P2 gene: developmental expression and homology of 5'-flanking sequences among fat cell-specific genes, Proceedings of the National Academy of Sciences, 83(11), 3786-90. Ingalls, A, M., Dıckie M. M. and Snell G. D., 1950, Obese, a new mutation in the house mouse, Journal of Heredity, 41(12), 317-8. Jocken, J. W., Langin, D., Smit, E., Saris, W. H., Valle, C., Hul, G. B., Holm, C., Arner, P. Blaak, E. E., 2007, Adipose triglyceride lipase and hormone-sensitive lipase protein expression is decreased in the obese insulin-resistant state, The Journal of Clinical Endocrinology & Metabolism, 92(6), 2292-9. Kane ,R., Godson, C. O'Brien, C., 2008, Chordin-like 1, a bone morphogenetic protein4 antagonist, is upregulated by hypoxia in human retinal pericytes and plays a role in regulating angiogenesis, Molecular Vision, 14, 1138-48. Kaya, A., 1996, Kırmızı et, Ege üniversitesi tarımsal uygulama ve araĢtırma merkezi, Teknik Bülten, 28. Kershaw, E. E. and Flier, J. S., 2004, Adipose Tissue as an Endocrine Organ, Journal of Clinical Endocrinology and Metabolism, 89(6), 2548–2556. Kershaw, E. E., Schupp, M., Guan, H. P., Gardner, N. P., Lazar, M. A. Flier, J. S., 2007, PPARgamma regulates adipose triglyceride lipase in adipocytes in vitro and in vivo, Endocrinology and Metabolism, American Journal of Physiology, 293(6), E1736-45. 56 Kim, H. B., Kong, M., Kim, T. M., Suh, Y. H., Kim, W. H., Lim, J. H., Song, J. H., and Jung, M. H., 2006, NFATc4 and ATF3 negatively regulate adiponectin gene expression in 3T3-L1 adipocytes, Diabetes, 55:1342-52. Komisarek, J. and Antkowiak, I., 2007, The relationship between leptin gene polymorphisms and reproductive traits in Jersey cows, Polish Journal of Veterinary Sciences, 10(4),193-7. Kosteli, A., Sugaru, E., Haemmerle, G., Martin, J. F., Lei, J., Zechner, R., Jr, A. W. F., 2010, Weight loss and lipolysis promote a dynamic immune response in murine adipose tissue, The Journal of Clinical Investigation, 120(10):3466-3479. Koza, R. A., Nikonova, L., Hogan, J., Rim, J. S., Mendoza, T., Faulk, C., Skaf, J. Kozak, L. P., 2006, Changes in gene expression foreshadow diet-induced obesity in genetically identical mice, PLOS Genetics, 2(5), e81. Kulig, H. Kmieć M., 2009, Association between leptin gene polymorphisms and growth traits in Limousin cattle, Genetika, 45(6), 838-41. Kumar, R. M., 2009, The Widely Used Diagnostics “DNA Microarray”-A Review, American Journal of Infectious Diseases 5(3), 214-225. Larman, B. W., Karolak, M. J., Adams, D. C. Oxburgh, L., 2009, Chordin-like 1 and twisted gastrulation 1 regulate BMP signaling following kidney injury, Journal of the American Society of Nephrology, 20(5),1020-31. Lagonigro, R., Wiener, P., Pilla, F., Woolliams, J. A. Williams, J. L., 2003, A new mutation in the coding region of the bovine leptin gene associated with feed intake, Animal Genetics, 34(5), 371-4. Larrain, J., Bachiller, D., Lu, B., Agius, E., Piccolo, S. De Robertis E. M., 2000, BMP binding modules in chordin: a model for signalling regulation in the extracellular space, Development, 127(4), 821-30. Lee, S. H., Cho, Y. M., Kim, B., Kim, N., Choy, Y., Kim, K., Yoon, D., Im, S., Oh, S. Park, E., 2008, Identification of marbling-related candidate genes in M. longissimus dorsi of high and low marbled Hanwoo (Korean Native Cattle) steers, BMB rep, 41, 846. Lee, K., Shin, J., Latshaw, J. D., Suh, Y. Serr, J., 2009, Cloning of adipose triglyceride lipase in poultry and expression of adipose triglyceride lipase during development of adipose in chickens, Poultry. Science, 88, 620-630. Lefterova, M. I. and Lazar, M. A., 2009, New developments in adipogenesis, Trends in Endocrinology and Metabolism, 20 (3), 108-114. Li, W. J., Li, H. B., Chen, J. L., Zhao, G. P., Zheng, M. Wen Q. J., 2008, Gene expression of Heart and Adipocyte Fatty Acid Binding Protein and correlation with intramuscular fat in Chinese Chickens, Animal Biotechnology, 19, 190-194. 57 Li, B., Shin, J. and Lee, K., 2009, Interferon-Stimulated Gene ISG12b1 Inhibits Adipogenic Differentiation and Mitochondrial Biogenesis in 3T3-L1 Cells, Endocrinology, 150, 1217–1224. Lieberman, M. and Marks, A. D., 2009, Basic medical biochemistry a clınical approach third edition, http://books.google.com.tr/books, [Ziyaret Tarihi: 12 Mayıs 2013]. Liefers, S. C., Veerkamp, R. F., Te Pas M. F., Delavaud, C., Chilliard, Y. Van Der Lende T., 2003, Leptin concentrations in relation to energy balance, milk yield, intake, live weight, and estrus in dairy cows, Journal of Dairy Science, 86(3), 799-807. Linford, N. J., Beyer, R. P., Gollahon, K., Krajcik, R. A., Malloy, V. L., Demas, V., Burmer, G, C. Rabinovitch, P. S, 2007, Transcriptional response to aging and caloric restriction in heart and adipose tissue, Aging Cell, 6(5), 673-88. Livak, K. J. and Schmittgen, T. D., 2001, Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT method, Methods, 25, 402-408. Lusk, J. L., 2007, Association of single nucleotide polymorphisms in the leptin gene with body weight and backfat growth curve parameters for beef cattle, Journal of Animal Science, 85, 1865-1872. Ma, S., Liui L., Li, Y. Ren, Y., 2009, Study on PPARgamma and aP2 mRNA expression of adipose tissues in obesity and obesity resistant rats, Wei Sheng Yan Jiu, 38(2), 163-5. Maury, E. and Brichard, S. M., 2010, Adipokine dysregulation, adipose tissue inflammation and metabolic syndrome, Molecular and Cellular Endocrinology 314, 1–16. Mersmann, H. J. and Smith, S. B., 2004, Adipose Tissue Development, Encyclopedia of Meat Sciences, 530-538. Millo, M. G., 2004, Adipose tissue and adipokines: for better or worse, Diabetes Metabolism, 30, 13-19. Nakayama, N., Han, C. E., Scully, S., Nishinakamura, R., He, C., Zeni, L., Yamane, H., Chang, D., Yu, D., Yokota, T. Wen D., 2001, A novel chordin-like protein inhibitor for bone morphogenetic proteins expressed preferentially in mesenchymal cell lineages, Developmental Biology, 232(2), 372-87. Narukami, T., Sasazaki, S., Oyama, K., Nogi, T., Taniguchi, M. and Mannen, H., 2011, Effect of DNA polymorphisms related to fatty acid composition in adipose tissue of Holstein cattle, Journal of Animal Science, 82 (3), 406-11. Ntambi, J. M. and Young-Cheul, K, 2000, Adipocyte differentiation and gene expression, Journal of Nutrition, 130(12), 3122-3126. 58 Ockner, R. K., Manning, J, A., Poppenhausen R. B. and Ho WK, 1972, A binding protein for fatty acids in cytosol of intestinal mucosa, liver, myocardium, and other tissues, Science, 177, 56-58. Oh, S. A., Suh, Y. M., Pang, G. Lee, K., 2011, Cloning of avian G(0)/G(1) switch gene 2 (G0S2) genes and developmental and nutritional regulation of G0S2 in chicken adipose tissue, Journal of Animal Science, 89, 367-375. Osuga, J. I., Ishibashi, S., Oka, T., Yagyu, H., Tozawa, R., Fujimoto, A., Shionoiri, F., Yahagi, N., Kraemer, F. B., Tsutsumi, O. and Yamada, N., 2000, Targeted disruption of hormone-sensitive lipase results in male sterility and adipocyte hypertrophy, but not in obesity, Proceedings of the National Academy of Sciences, 97, 787-792. Öztürkcan, O., Demir E. ve Görgülü M 1996, Çiftlik Hayvanlarında Yaglanma, Çukurova Üniversitesi Ziraat Fakültesi Genel Yayın No:136, Yardımcı Ders Kitapları Yayın No:12, Adana. Petrovic, N., Shabalina, I. G., Timmons, J. A., Cannon, B. Nedergaard, J., 2008, Thermogenically competent nonadrenergic recruitment in brown preadipocytes by a PPARgamma agonist, Endocrinology and Metabolism, American Journal of Physiology, 295(2), E287-96. Piccolo, S., Sasai, Y., Lu, B. De Robertis E. M., 1996, Dorsoventral patterning in Xenopus: inhibition of ventral signals by direct binding of chordin to BMP-4, Cell, 86(4), 589-98. Piya, M. K., McTernan, P. G. and Kumar, S., 2013, Adipokine inflammation and insulin resistance: the role of glucose, lipidsand endotoxin, Journal of Endocrinology, 216 (1), T1-T15. Puente, B. A., Feve, B., Fellahi, S. Bastard, J. P., 2008, Adipokines: The missing link between insulin resistance and obesity, Diabetes and Metabolism, 34, 2-11. Rosen, E. D., Walkey, C. J., Puigserver, P. and Spiegelman, B. M., 2000, Transcriptional regulation of adipogenesis, Genes and Development, 14, 12931307. Sakuta, H., Suzuki, R., Takahashi, H., Kato, A., Shintani, T., Iemura, Si., Yamamoto, T. S., Ueno, N. Noda, M., 2001, Ventroptin: a BMP-4 antagonist expressed in a double-gradient pattern in the retina, Science, 293(5527), 111-5. Samulin, J., Berget, I., Lien, S. Sundvold, H., 2008, Differential gene expression of fatty acid binding proteins during porcine adipogenesis, Comparative Biochemistry and Physiology Part B, Biochemistry and Molecular Biology, 151(2), 147-52. Schena, M., Shalon, D., Davis R. W. and Brown P. O., 1995, Quantitative Monitoring of Gene Expression Patterns with a Complementary DNA Microarray, Science, 270 (5235), 467-470. 59 Schenkel, F. S., Miller, S. P., Ye, X., Moore, S. S., Nkrumah, J. D., Li, C., Yu, J., Mandell, I. B., Wilton, J. W. Williams, J. L., 2005, Association of single nucleotide polymorphisms in the leptin gene with carcass and meat quality traits of beef cattle, Journal of Animal Science, 83(9), 2009-20. Seale, P., Kajimura, S., Yang, W., Chin, S., Rohas, L. M., Uldry, M., Tavernier, G., Langin, D. Spiegelman, B. M., 2007, Transcriptional control of brown fat determination by PRDM16, Cell Metabolism, 6(1), 38-54. Serr, J., Suh, Y. and Lee, K., 2009, Regulation of adipose triglyceride lipase by fasting and refeeding in avian species, Poultry Science, 88, 2585-91. Serr J., Suh, Y. S., Oh, Shin, A. S., Kim, M. S., Latshaw, J. D. and Lee, K., 2011, Acute up-regulation of adipose triglyceride lipase and release of non-esterified fatty acids by dexamethasone in chicken adipose tissue, Lipids, 46, 813-820. Shan T., Wang, Y., Wu, T., Guo, J., Liu, J., Feng, J. Xu, Z., 2008. Porcine adipose triglyceride lipase complementary deoxyribonucleic acid clone, expression pattern, and regulation by resveratrol, Journal of Animal Science, 86, 1781-1788. Shin, J., Lim, S., Latshaw, J. D. Lee, K., 2008, Cloning and expression of delta-like protein 1 messenger ribonucleic acid during development of adipose and muscle tissues in chickens, Poultry Science, 87(12), 2636-46. Shin, J., Li, B., Davis, M. E., Suh, Y. Lee, K., 2009, Comparative analysis of fatty acidbinding protein 4 promoters: conservation of peroxisome proliferator-activated receptor binding sites, Journal of Animal Science, 87(12), 3923-34. Spiegelman, B. M., Frank, M. Green, H., 1983, Molecular cloning of mRNA from 3T3 adipocytes. Regulation of mRNA content for glycerophosphate dehydrogenase and other differentiation-dependent proteins during adipocyte development, The Journal of Biological Chemistry, 258(16), 10083-9. Steinberg, G. R., Kemp, B. E. Watt, M. J., 2007, Adipocyte triglyceride lipase expression in human obesity, Endocrinology and Metabolism, American Journal of Physiology, 293(4), E958-64. Stock, M. J. and Cinti, S., 2003, Structure and Function of Brown Adipose Tissue, Encyclopedia of Food Sciences and Nutrition (Second Edition), 29-34. Su, A. I., Wiltshire, T., Batalov, S., Lapp, H., Ching, K. A., Block, D., Zhang, J., Soden, R., Hayakawa, M., Kreiman, G., Cooke, M. P, Walker, J. R. Hogenesch, J. B., 2004, A gene atlas of the mouse and human protein encoding transcriptomes, Proceedings of the National Academy of Sciences, 101(16), 6062-7. 60 Takahashi, H., Shintani, T., Sakuta, H. Noda, M., 2003, CBF1 controls the retinotectal topographical map along the anteroposterior axis through multiple mechanisms, Development, 130(21),5203-15. Taniguchi, M., Guan, L. L., Zhang, B., Dodson, M. V., Okine, E., Moore, S. S., 2008, Gene expression patterns of bovine perimuscular preadipocytes during adipogenesis, Biochemical and Biophysical Research Communications, 366, 346351. Tchkonia, T., Lenburg, M., Thomou, T., Giorgadze, N., Frampton, G., Pirtskhalava, T., Cartwright, A., Cartwright, M., Flanagan, J., Karagiannides, I., Gerry, N., Forse, R. A., Tchoukalova, Y., Jensen, M. D., Pothoulakis, C. Kirkland, J. L., 2007, Identification of depot-specific human fat cell progenitors through distinct expression profiles and developmental gene patterns, Endocrinology and Metabolism: American Journal of Physiology, 292(1), E298-307. Thorrez, L., Van Deun, K., Tranchevent, L. C., Van Lommel, L., Engelen, K., Marchal, K., Moreau, Y., Van Mechelen, I. Schuit, F., 2008, Using ribosomal protein genes as reference: a tale of caution, PLoS One, 3(3), e1854. Türkmen, Y. E., 2003, Esansiyel Amino Asitler, www.kimyasanal.net/konugoster.php [Ziyaret Tarihi: 01 ġubat 2013]. Umucalılar, H. D. ve GülĢen N., 2005, Çiftlik Hayvanlarında Beslenme Hastalıkları. S.Ü. Basımevi, Konya. Wang H. B., Li, H., Wang, Q. G., Zhang, X. Y., Wang, S. Z., Wang, Y. X., Wang, X. P., Wang, S. P., Laurin, N., Himms Hagen, J., Rudnicki, M. A., Levy, E., Robert, M. F., Pan, Oligny, L. L. and Mitchell, G. A., 2001, The adipose tissue phenotype of hormone-sensitive lipase deficiency in mice, Obesity Research, 9, 119-128. Wang, Y. H., Byrne, K. B., Reverter, A., Harper, G. S., Taniguchi, M., McWilliam, S. M., Manen, H., Oyama, K. and Lehnert, S. A., 2005, Transcriptional profiling of skeletal muscle tissue from two breeds of cattle, Mammalian Genome, 16(3), 201210. Webber, J., 2003, Energy balance in obesity, Proceedings of the Nutrition Society, 62(2), 539-43. Woronuk, G. N., Marquess, F. L., James, S. T., Palmer, J., Berryere, T., Deobald, H., Howie, S. Kononoff, P. J., 2012, Association of leptin genotypes with beef cattle characteristics, Animal Genetics, 43(5), 608-10. Wylie, A. and Newforge, A. F. B. I., 2007, Leptin where’s the beef ?, http://www.afbini.gov.uk [Ziyaret Tarihi: 10 Nisan 2013]. Xu, Q. L., Tang, G. W., Zhang, Q. L., Huang, Y. K., Liu, Y. X., Quan, K., Zhu, K. Y. Zhang, C. X., 2011, The FABP4 gene polymorphism is associated with meat tenderness in three Chinese native sheep breeds, Czech Journal of Animal Science, 1, 1-6. 61 Yamasaki, T., Tahara, K., Takano, S., Murayama, M. I., Rose, M. T., Minashima, T., Aso, H. and Ito, S., 2006, Mechanism of plasma glutathione peroxidase production in bovine adipocytes, Cell and Tissue Research, 326, 139-147. Yarru, L, P., 2008, Effects of Aflatoxin on Hepatic Gene Expression in A Poultry Model, Faculty of the Graduate School University of Missouri Columbia, Master Thesis. Zhang, Y, Y., Zan, L. and Wang, H., 2010, Screening candidate genes related to tenderness trait in Qinchuan cattle by genome array, Molecular Biology Reports, 38(3), 2007-14. Zheng, Z., Yin, Z., Gao, W., Zhang, H., Hu, Y. Zhang, S., 2010, Effects of neurogenesin 1 gene on functional recovery of spinal cord injury in rats and its mechanism, Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi, 24(4), 410-4. Zimmerman, L. B., De Jesús Escobar, J. M. Harland, R. M., 1996, The Spemann organizer signal noggin binds and inactivates bone morphogenetic protein 4, Cell, 86(4), 599-606. Zimmerman A.W. and Veerkamp J. H., 2002, New insights into the structure and function of fatty acid-binding proteins, Cellular and Molecular Life Sciences, 59, 1096-1116. Zimmermann, R., Strauss, J. G., Haemmerle, G., Schoiswohl, G., R. Gruenberger, B., Riederer, M., Lass, A., Neuberger, G., Eisenhaber, F., Hermetter, A. Zechner, R., 2004. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase, Science, 306, 1383-1386. ZoBell D. R., Whittier, D. and Holmgren, L., 2005, Beef quality and yield grading, extension.usu.edu/cyberlivestock/.../beef3.pdf [Ziyaret Tarihi: 05 Haziran 2013]. 62 ÖZGEÇMĠġ KĠġĠSEL BĠLGĠLER Adı Soyadı Uyruğu Doğum Yeri ve Tarihi Telefon Faks e-mail : : : : : : Selçuk KAPLAN Türkiye Cumhuriyeti Erzurum/Merkez- 12.09.1979 0 332 223 36 37 0 332 241 01 08 [email protected] EĞĠTĠM Derece Lise : Üniversite : Yüksek Lisans : Doktora : Adı, Ġlçe, Ġl Atatürk Lisesi, Kırklareli Adnan Menderes Üniversitesi, Aydın Selçuk Üniversitesi, Konya Selçuk Üniversitesi, Konya Bitirme Yılı 1997 2005 2010 Devam ediyor Ġġ DENEYĠMLERĠ Yıl 2008 Kurum S.Ü. Ziraat Fakültesi Görevi AraĢtırma Görevlisi YABANCI DĠLLER: Ġngilizce YAYINLAR Kaplan, S. and Boztepe, S., 2010, The Determination of Prolactin Gene Polymorphism Using PCR-RFLP Method within Indigenous AnatolianWater Buffalo and Brown Swiss, 2nd International Symposium on SustainableDevelopment. 8-9 June 2010, Saraybosna (Yüksek Lisans tezinden hazırlanmıĢtır). Aytekin, Ġ. ve Kaplan, S., 2010, Koyunlarda vücut kondüsyon puanlaması ve yetiĢtiricilikte önemi, HasadHayvancılık, 303, 42-45. Kaplan, S. ve Boztepe S., 2011, Çiftlik Hayvanlarında Refah, Konya Ticaret Borsası Yayını, Adım Matbacılık, Konya.