DNA_Replikasyonu_RNA_Transkripsiyonu_Protein_Sentezi400
advertisement

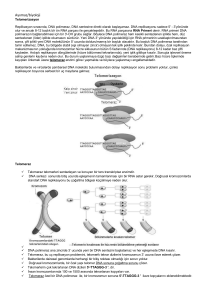
DNA REPLİKASYONU Prof.Dr. Hüseyin SÖNMEZ Cerrahpaşa Tıp Fakültesi Biyokimya Anabilim Dalı • DNA genetik bilginin depolandığı biyolojik makro • • • moleküldür. Prokaryotlarda hücrenin diğer komponentleri ile birlikte bulunur. Ökaryotlarda ise nükleus içinde yer alır ve nükleus zarı ile diğer hücresel komponentlerden ayrılmıştır. Ökaryotlarda DNA proteinlere bağlı olarak kromatin adı verilen yapılar oluşturur. • DNA çok sayıda deoksiribo nükleotid birimlerinin • • • birleşmesinden oluşur. Nükleotidler azotlu baz, pentoz şekeri ve fosfat grubu olmak üzere 3 grubu içerir. Azotlu bazlar pürin ve pirimidin olarak 2 tiptir. Pentoz şekeri ise deoksiribozdur • DNA’nın 2 polinükleotid zincirinden oluşan sarmal heliks yapısı 1953’de Watson ve Crick tarafından ortaya çıkarıldı. • Heliks yapısında yer alan 2 polinükleotid zincirin bazları arasında oluşan hidrojen bağları ile birleşir. • Bir merkez eksen etrafında kıvrılarak ikili sarmal yapıyı meydana getirir.Polinükleotid zincirinde yer alan 2 nükleotid arasında fosfodiester bağı vardır. • Her baz çiftinde bir zincirdeki bir pürin diğer zincirdeki pirimidin ile hidrojen bağları aracılığı ile birleşir. • Adenin-timin arasında 2, guanin-sitozin arasında 3 hidrojen bağı oluşur. • Her iki zincirin bazları düzlemsel yapıdadır ve düzelemleri eksene diktir. • Heliks sarmalın bir tam dönüşünde 10 baz çifti yer alır. • DNA’nın birbirini tamamlayan iki komplementer zinciri zıt yönlü yani anti paraleldir. Bir zincir 5’ Æ3’ yönünde iken diğeri 3’ Æ 5’ yönündedir. DNA Replikasyonu’nun Genel Özellikleri • DNA replikasyonu semikonservatiftir. • DNA’nın her bir ipliği yeni ipliğin sentezi için bir kalıp fonksiyonu görür. • Bu şekilde üretilen iki yeni DNA molekülünde bir yeni sentez edilen iplik bir de eski iplik yer alır. • Replikasyonun semi konservatif olduğu Meselson-Stahl deneyi ile gösterilmiştir. • DNA replikasyonunda polimeraz enzimleri dışında 20’den fazla enzim, protein görev alır. Tümüne DNA replikaz sistemi veya Replizom denir. • DNA replikasyon aşamaları Başlama safhası – Replikasyon orjini (leri)’nin belirlenmesi – Çift iplikli DNA’nın tek iplikli yapı oluşturmak üzere denatürasyonu – Replikasyon çatalı oluşumu Uzama safhası Sonlanma safhası Başlama safhası • DNA polimeraz kalıp olarak sadece tek zincirli DNA molekülü kullanır. • Çift sarmal DNA molekülünün replikasyon öncesinde birbirinden ayrılması gerekir. • DNA replikasyonu prokaryotlarda replikasyon orijini olarak adlandırılan özel bir noktada başlar. • Bu noktada replikasyon çatalı oluşur. • DNA epilasyonu ökaryotlarda prokaryotların aksine birçok noktada birden başlar. • Bu şekilde prokaryot DNA’sına göre çok daha büyük olan ökaryot DNA’sıın replikasyonu kısa sürede tamamlanır. Prokaryotlarda DNA Replikasyonu • E.coli’de DNA’sı çift iplikli halkasal yapıdadır. • DNA replikasyonu Ori C olarak adlandırılan replikasyon merkezinde başlar.Bu merkezin özelliği tekrar eden 2 anahtar dizi içermesidir. • Birinci dizi 9 baz çiftinin 4 kez tekrar edilmesi, ikinci dizi 13 baz çiftinin 3 kez tekrar edilmesidir. • Replikasyon merkezinde replikasyonun başlayabilmesi için DNA’nın iki zincirinin birbirinden ayrılması ve replikasyon çatalının meydana gelmesi gerekir. • Bu amaç için farklı protein ve enzimler fonksiyon görür. • Başlama safhasındaki anahtar komponent DnaA proteinidir. • 20 civarında DnaA protein molekülünden oluşan bir kompleks merkezde tekrar eden 9 baz çiftinin bulunduğu bölgeye bağlanır. Bu işlemde ATP harcanır. • 13 baz çiftinin bulunduğu bölgede DNA denatüre edilir. • Bu işlemde ATP ve HU olarak adlandırılan histon benzeri protein kullanılır. • Takiben DnaB proteini, DnaC proteinini içeren bir reaksiyon • • • • ile denatüre olan bölgeye bağlanır. DnaB helikaz olarak etki eder ve replikasyon çatalı oluşturur. Ayrılmış olan DnaA zincirlerinin her biri “tek zincirli DNA bağlayıcı protein” olarak adlandırılan SSB proteinleri ile etkileşir (single stranded binding protein) Bu proteinler tek zincirli DNA’yı stabilize eder ve renatürasyonu önler. Çift sarmalın açılması ile oluşan topolojik gerilim topoizomeraz (DNA gyrase) tarafından dengelenir. Uzama safhası • Bu safhada replikasyon çatalı ile oluşan iki ana zincir yeni DNA iplikçiklerinin sentezi için kalıp görevi görür. • Ana zincirleri kalıp olarak kullanıp yeni DNA sentezini yapacak olan enzim DNA polimerazdır. • DNA polimeraz bu sentezi 5’Æ3’ yönünde yapabilir. • E.coli’de 3 tip DNA polimeraz mevcuttur. I, II ve III tipleri • DNA replikasyonu polimeraz III tarafından yapılır. • DNA polimerazın substratları dört deoksiribonükleotid trifosfattır. dATP, dCTP, dGTP, dTTP • DNA polimeraz iki serbest nükleotid arasındaki reaksiyonu kataliz edemez. • Polimeraz ancak mevcut bir kalıbın serbest 3’OH ucuna nukleotid ilavesi yapabilir. Bu ön zincir Primer olarak adlandırılır. • Primer DNA polimeraz için bir nukleotidi ilk kabul edici yer olarak görev yapar. • Primer, primaz olarak adlandırılan bir enzim tarafından sentez edilen ve birkaç nukleotidden oluşan bir RNA parçasıdır. DNA kalıbı T C G A G C T A 3’ OH Primer C G G C G G dG TP ÆPPi OH • DNA Replikasyonu DNA molekülünün ayrılan iki zincirinde aynı anda meydana gelir, ancak aynı şekilde cereyan etmez. • Bir zincir replikasyon çatalına doğru kesintisiz olarak kopya edilebilir. • Diğer zincir ise kısa fragmentler şeklinde sentez edilir ve bu parçalar daha sonra birleştirilir. Bunlara Okazaki fragmentleri denir. • Bir okazaki fragmenti tamamlandığı zaman RNA primer polimeraz 1 tarafından uzaklaştırılır. Arada kalan boşluklar DNA ligaz ile doldurulur. • Ligaz bir iplikçiğin 3’ hidroksil ucu ile diğerinin 5’ fosfat ucu arasında bir fosfodiester bağının oluşumunu katalizler. Sonlanma safhası • E.coli’de DNA’nın iki replikasyon çatalı terminal bölgede birleşir. • Bu bölge 20 baz çiftinden oluşur ve Ter. adını alır. • Bu bölge aynı zamanda TUS (terminus utilization substance) adlı protein için bağlanma bölgesi içerir ve replikasyon bu bölgede durur. Ökaryotlarda DNA Replikasyonu • Ökaryotik hücrenin siklusu 4 faz içerir Ökaryot DNA replikasyonu • Ökaryot hücrelerde DNA replikasyonundaki en belirgin fark birçok replikasyon orjini içermesidir. • Ökaryot hücrelerde DNA replikasyonu hücre döngüsünün S fazında cereyan eder. • Replikasyon orjinlerine ARS (Autonomusly replicating sequences) denir. • S fazı öncesi G1 fazı esnasında bütün ARS dizilerine bazı özel proteinler bağlanır ve orijin tanı kompleksleri ÖRL’ler (origin recognition complex) oluşur. • S fazında özel kinazlar ORC’ye bağlanarak DNA polimerazın hızlanmasına olanak sağlayan ön replikasyon komplekslerini oluşturur. Ökaryot DNA polimerazlar α δ Ε β γ Nükleer DNA replikasyonu Tamir fonksiyonu Mitokondrial DNA replikasyonu • Ökaryotik kromozomlar prokaryotik kromozomlardan farklı olarak doğrusal yapıdadır. • Kromozomların ucu telomer olarak adlandırılan yapılar ile sonlanır. • Bu durum DNA replikasyonunun sonlanmasında kesintili zincirde problem yaratır. • Hücreler normalde yaklaşık 50 kez bölünürler (Hayflick limiti) • Bu süreçte telomerler progressif olarak kısalır • Her hücre bölünmesinde kısalan telomer kritik uzunluğa gelince hücre bölünmesi durur Æ Yaşlanma ve Ölüm • Telomeraz denilen enzim aktif olursa hücre büyümeye ve bölünmeye devam eder TELOMERAZ • Ribonukleoprotein yapıda bir revers transkriptazdır • Kromozom uçlarına telomerik tekrarların eklenmesini katalizler TELOMERAZ • Bazı fetal dokular, germ hücreleri, kök hücre, replikatif kapasitesi yüksek bazı hücreler (sperm, ovum, epidermal deri hücresi vb.) ve kanser hücreleri telomeraz aktivitesi gösterir • Somatik hücrelerde ise yok veya ölçülemeyecek kadar düşüktür • Telomeraz aktivitesi çocuklarda erişkinlere göre daha yüksektir DNA (Kromozom) Mutasyonları • Mutasyonlar başlıca 2 şekilde görülebilir. 1-Kromozom sayısında meydana gelen değişimler 2-DNA’nın nükleotid dizisinde meydana gelen değişiklikler 1-Kromozom sayısında meydana gelen değişimler Örneğin: -Diploid genomdan tek kromozom kaybı: Monozami (2n-1) -Diploid genoma tek kromozom ilavesi *Trizomi (2n+1) *Down Sendromu (Kromozom 21) 2-DNA’nın nükleotid dizisinde meydana gelen değişiklikler -DNA üzerindeki bu tip mutasyonlara DNA replikasyonu ya da onarımı sırasındaki hatalar, fiziksel ve kimyasal etkenler yol açar. 1-Nokta mutasyonları Transisyon Transversiyon 2-Çerçeve kayması mutasyonları Delesyon İnsersiyon Nokta Mutasyonları • En çok rastlanan tiptir. Tek bir baz çiftinin değişimi ile meydana gelir. Transisyon Tip Bir pürin diğer pürin veya bir Pirimidin diğer bir pirimidin ile yer değiştirir O6 GGG Æ AGG Glisin Arginin Transversiyon tip Pürin yerine pirimidin ya da Pirimidin yerine pürin katılır. • • Çerçeve Kayması Mutasyonları (Delesyon) DNA yapısına bir baz girmesi veya çıkması ile oluşur. Normal kodonlarda nükleotid dizisinin okuma çerçevesi değişir ve amino asid dizilimi kayar. Liz AAG Gln GAA Arj AGA C AAG Liz AAG AGA A Arg CAG GAA AGA G AAG AAG AA RNA Transkripsiyonu • DNA’nın yapısında deoksiribonükleotidlerin doğrusal dizilimi yer alır. • Bu dizi genlerin son ürünü olan proteinlerin yapımı için komut verir. • Bir DNA kalıbından RNA moleküllerinin sentezlenmesi işlemine transkripsiyon (okuma) adı verilmektedir. • Tüm ökaryotik hücreler 4 tip RNA içerir. Replikasyon ve Transkripsiyon arasındaki belirgin farklar • Replikasyon esnasında tüm DNA molekülü kopyalanır. • • • • • Transkripsiyonda ise her defasında yalnız özel gen ya da gen grupları transkribe edilir. Bazı DNA bölgeleri ise hiçbir zaman transkripsiyona uğramaz. Spesifik baz dizileri transkribe edilecek olan DNA segmentinin başlangıcını ve sonunu işaretler. Transkripsiyonda da başlama, uzama ve sonlanma safhaları vardır. Transkripsiyon bir primere ihtiyaç göstermez. DNA’nın yalnız bir iplikçiliği kalıp olarak fonksiyon görür. Transkripsiyon sonucu sentez edilen molekül ekleme, çıkarma gibi çeşitli modifikasyonlara uğrar ve fonksiyonel şekle dönüşür. Prokaryotik Gen Transkripsiyonu • DNA ikili sarmalının zincirlerinden biri üzerindeki bir bölgeye • • komplementer olan RNA molekülünün sentezlenmesi işlemi RNA polimeraz enzimi ile yapılır. Prokaryotik hücreler tek tip RNA polimeraz içerir. Bu RNA polimeraz farklı RNA tiplerini sentez edebilir. RNA polimeraz yaklaşık 500 kd büyüklüğünde ve birçok alt üniteden oluşan kompleks bir enzimdir. Halo enzim α2 β β’ w σ Çekirdek enzim Promotor bölgeyi tanır ve Sentezi başlatır. • RNA polimeraz aktivite için çift iplikli RNA’ya gereksinim duyar ve bir iplik kalıp olarak kullanılır. • RNA sentezi, RNA polimeraz emiminin DNA molekülü üzerinde promotor olarak adlandırılan spesifik baz dizilerini tanıması ile başlar. • Promotor diziler transkripsiyonun başlama bölgesini belirler. Bu dizilerdeki mutasyonlar genin ifade edilmesini ciddi biçimde etkiler. Bakteriyel promotorlarda konsensus diziler 1-Transkripsiyon başlama noktasının 10 nükleotid uzaktaki-10 bölgesi Pribnow kutusu ÆTATAAT 2-Başlama noktasına 35 nükleotid uzaktaki –35 bölgesi TGTTGACA • DNA üzerindeki promotor bölgeyi tanıyan RNA polimeraz üzerindeki sigma alt birimidir. • Bu alt birim enzimin core kısmına geçici olarak bağlanır ve enzimi spesifik bölgeye yönlendirir. • Sigma alt birimi sentezin başlamasını takip eden birkaç fosfodiester bağından sonra enzimin core kısmından ayrılır. • RNA polimeraz enziminin bağlanması DNA çift iplikçiğinin açılmasına ve transkripsiyon kabarcığının oluşumuna neden olur. Yaklaşık 17 bazlık bir bölge açılır. • RNA polimeraz etkisi ile RNA polimerizasyonu ortamdaki ribonükleotidlerin fosfodiester bağı ile birbirlerine bağlanması şeklinde devam eder. • Sentez 5’Æ3’ yönündedir. • E.coli’de sentez hızı yaklaşık 50 nükleotid/sn’dir. Sonlanma • Transkripsiyonun sonlanma safhası da hassas bir kontrol altındadır. • Kalıp olarak kullanılan DNA iplikçiğinin transkribe edilen bölgeleri sonlanma sinyalleri içerir. • Bakterilerde iki tür sonlanma mevcuttur. 1-p (rho) faktörünün gerekli olduğu sonlanma 2-rho faktöründen bağımsız sonlanma • Rho faktörü bir hekzamerik proteindir ve sonlanma bölgesinde tek zincirli RNA’ya bağlanarak kalıp DNA’dan ayrılmasını stimüle eder. • rho faktöründen bağımsız sonlanma da GC ve takiben AT bazlarından zengin olan sonlanma bölgesinde RNA transkripti saç tokası şeklinde bir yapı oluşturabilir ve polimeraz enzimi yavaşlar. Ökaryotik Gen Transkripsiyonu • Ökaryotlarda 3 tip RNA polimeraz vardır. α-amanitine karşı duyarlılığı Tip Lokalizasyon Hücresel transkriptleri I II II Nukleolus Nukleoplazma Nukleoplazma 18S, 5.8S ve 28S rRNA mRNA prekürsör ve hnRNA tRNA ve 5S rRNA Duyarsız Kuvvetli inhibe olur Yüksek konsantrasyonda duyarlı • Ökaryotlarda transkripsiyonun başlama bölgelerinde prokaryotlara benzer biçimde özel baz dizilimleri içeren bölgeler vardır. 1—30 bölgesinde bulunan TATA kutusu 2—80 bölgesinde yer alan CAAT kutusu • Rifampisin ve aktinomisin transkripsiyonu inhibe eden iki antibiyotiktir. • Rifampisin RNA sentezinin RNA zincirindeki ilk birkaç fosfodiester bağını oluşumunu engelleyerek transkripsiyonu inhibe eder. • Aktinomisin spesifik olarak çift iplikli DNA’ya bağlanarak RNA transkripsiyonunu inhibe eder. • Ayrıca amanita phalloides adlı mantar tarafından üretilen alfa amanitin RNA polimeraz II’ye bağlanarak RNA sentezinin uzama safhasını bloke eder. • Yüksek konsantrasyonlardaki alfa amanitin polimeraz III’ü inhibe edebilir buna karşılık polimeraz I bu toksine karşı duyarlı değildir. • Sentez edilen RNA molekülleri primer transkripsiyon adını alır ve modifikasyona uğrayarak olgun şekle dönüşür. • Olgun RNA molekülüne dönüşüm post transkripsiyonel modifikasyon olarak isimlendirilir. • Prokaryotik tRNA ve rRNA molekülleri ile tüm ökaryotik RNA molekülleri bu modifikasyona uğrar. mRNA Modifikasyonu 5’ cap oluşumu • Ökaryotlarda RNA polimerazII kalıp DNA zincirinden büyük bir primer transkript sentezler • • Sentez edilen bu primer transkriptin 5’ ucuna bir cap takılır. • • Bu difosfatın β fosfatı GTP’nin α fosfatı ile etkileşir. • Bu yapı molekülü nükleazların yıkımına karşı korur ve aynı zamanda olgun mRNA’nın ribozomlara bağlanmasında rol alır. 5’ cap oluşturmak için bu uçta bulunan nükleotidin terminal trifosfatı bir fosfatını kaybeder, 5’ difosfat oluşur. S adenozin metioninden (SAM) bir metil grup guanin halkanın 7.pozisyonuna transfer edilir. Böylece 7 metil guanizin cap yapısı oluşur. mRNA ‘ya PoliA zincirinin eklenmesi • PoliA zinciri çekirdek enzimi olan poliadenilat polimeraz ile oluşur. • AAUAAA dizisine yakın bir noktada nükleotidler ekzonukleaz ile uzaklaşır. Buraya yaklaşık 250 nukleotidlik poliA zinciri ilave edilir. Bu yapı mRNA’nın stabilize olmasını ve çekirdekten çıkışını kolaylaştırır. • Ökaryötik pre-mRNA transkripti ekzon ve intron adı verilen bölgeler içerir. Ekzonlar olgun mRNA’da mevcut iken, intronlar transkriptten uzaklaştırılır. Bu nedenle intronlar proteinin aminoasit dizisine katkıda bulunmaz. • Pre-mRNA transkriptindeki intron-ekzon ayrım bölgelerindeki diziler (5’) GU ve (3’) AG’dir. • Splays (kesip, çıkarma, birleştirme) mekanizmalarına göre intronlar çeşitli gruplara ayrılır. • Grup I ve Grup II intronlar herhangi bir ilave protein ya da enzimatik aktivite gereksinimi olmaksızın kesip çıkarma işlemini kendileri yapabilir (self splicing) • Splays mekanizması her iki grupta da benzer biçimde iki transesterifikasyon mekanizması içerir. • Grup I intronlar bir guanin nükleotid kofaktöre gereksinim duyar. Ancak bu enerji kaynağı olarak kullanılmaz. • Guaninin 3.hidroksil grubu bir nükleofil olarak kullanılır. • Grup II intronlardaki reaksiyon da Grup I’e benzer ancak bir guanin nükleotid kofaktör olarak kullanılmaz. • İntronların üçüncü ve en büyük grubu nüklear mRNA primer transkriptlerinde bulunur. • Bunlar splaysozomal intronlar olarak adlandırılır. • Bu intronların kendileri kesip çıkarma işlemini yapamaz.İlave olarak büyük protein komplekslere ihtiyaç duyar. Bu kompleks splaysozom olarak adlandırılır. Bu yapılar özelleşmiş RNA protein kompleksleri, küçük nüklear ribonükleoproteinler (snRNPs) içerir. • Her snRNPs’ler de küçük nüklear RNAs içerir (snRNAs) • Nüklear ribonükleoproteinler olan U1 ve U2 introna bağlanır. Oluşan kompleks splaysozom olarak adlandırılır. • U1 intron-ekzon bağlantı bölgesine bağlanırken U2 intron içerisinde adenin içeren bölgeye bağlanır. • Diğer ribonükleoproteinler U4,U5,U6 komplekse bağlanır ve bir loop oluşturulur. • İntronun 5’ ucundaki G kalıntısına bağlı fosfat grubu adenin kalıntısının 2’ hidroksil grubu ile bağ oluşturur. • Bir yıkım meydana gelir. • İntronun 3’ ucunda ikinci yıkım oluşur. Ekzonlar birleştirilir. Ökaryotik rRNA Modifikasyonu • rRNA geni 45S büyüklüğünde bir transkript oluşturur. • Bu transkript proteinler ile birleşerek ribonükleoprotein partikülleri oluşturur. • 45S transkriptinin nükleotidlerinin %1-2’si metillidir. Bu metik grupları transkriptin yıkımı çin marker fonksiyonu görür. • Meydana gelen bir seri yıkım sonucu 18S, 28S ve 5.8S rRNA’lar meydana gelir. Ökaryotik tRNA Modifikasyonu • tRNA primer transkripti yaklaşık 100 nükleotid • • • uzunluğunda oluşturulur. Bu primer transkript 5’ ve 3’ uçlarından yıkılarak intronlar uzaklaştırılır. Bu olayda endonükleazlar görev yapar. Daha sonra 3 ayrı modikasyon meydana gelir. 1-Urasil SAM tarafından metillenir. Timin oluşur 2-Urasilin çift bağlarından biri dihidrourasil oluşturmak üzere indirgenir 3-Bir urasil kalıntısından pseudoüridin oluşturulur. Son olarak 3’ uca CCA dizisi ilave edilir. Genetik Şifre ve Protein Sentezi • Protein sentezi biyosentetik işlemler arasında en kompleks olanlardandır. • Protein sentezi çok sayıda ribozomal proteinleri, enzimleri, protein faktörleri, tRNA’ları içerir. • Bir hücrenin tüm biyosentetik reaksiyonlar için kullandığı kimyasal enerjinin ∼ %90’ı protein sentezi için harcanır. • Protein sentezinin bu çok kompleks yapısına karşın hızı oldukça yüksektir. E.coli’de 100 amino asidlik bir polipeptidin sentezi ∼ 5 sn. sürer. • DNA’nın yapısındaki deoksiribonükleotidlerin doğrusal dizilimi genlerin son ürünü olan proteinlerin yapımı için komut verir. • 1961’de Francois Jacob ve Jacques Monod mRNA’nın varlığını ortaya atmış ve bu buluş genetik bilginin DNA’da depolandığı halde, proteine çevrilen şifrenin RNA’da bulunduğuna açıklık getirmiştir. • Asıl sorun dört harfin (Dört nükleotidin) 20 amino asidin dizilimini nasıl belirlediğidir. • 20 tür amino asidin protein molekülündeki diziliş sırasının belirlenmesi için azotlu bazlarında özel bir diziliş göstermesi gerekir. • Bu ancak özel bir kodlama sistemi ile olabilir. • Bu kodlama sisteminde her aminoasid için bir baz olsa 41=4 kod olur (4<20). • Bu kodlama sisteminde her aminoasid için iki baz olsa 42=16 kod olur (16<20). • Her amino asid için 3 bazdan oluşan bir kod olur ise 43=64 kod olur (64>20). Genetik şifre (Kod)’nin genel özellikleri • Genetik şifre mRNA’daki ribonükleotid bazları kullanılarak doğrusal olarak yazılır. • mRNA’daki üç ribonükleotid bir kodon’u oluşturur ve her bir kodon bir amino asidi belirler. • Genetik şifre dejeneredir. • Bir amino asid birden fazla kodon tarafından belirlenebilir. 18 aminoasidin birden fazla kodonu vardır. • Aynı amino asidi belirleyen kodonlara sinonim kodonlar denir. • Şifrede başla ve dur sinyalleri bulunur. – AUG=Başlama kodonu – UAA, UAG,UGA=Sonlanma kodonları • Şifre içinde virgüller yoktur. Şifre duraksamazdır (Kodonlar arasında bir boşluk yoktur). • Şifre evrenseldir (virüs, probaryat, ökaryotlar aynı şifreyi kullanır). Ancak bazı istisnalar saptanmıştır. • mRNA’nın translasyonu amino asidlerin polipeptid zincirine biyolojik polimerizasyonudur. • Bu işlem ribozomlarda gerçekleşir. Ribozomal Yapı • 150-200 Ao büyüklüğünde partiküllerdir. Sitoplazmada ya serbest halde ya da endoplazmik retikulumun yüzüne yapışık olarak bulunurlar. • • • • Yapısal olarak rRNA ve protein içerirler. Bir büyük bir küçük olmak üzere iki alt birim içerirler. Alt üniteler Svedberg birimi ile tanımlanır. Ultra santrifüjde oluşan çekim alanında hareket eden cisimciklerin çökelme hızlarını belirleyen bir birim olup, cisimciğin kütlesi ile ilişkilidir. • mRNA’nın translasyonu ile ilgili başlıca sorulardan biri, sitozoldeki aminoasidlerin mRNA’daki uygun kodonlarına nasıl doğru olarak aktarıldığıdır. • tRNA’nın bulunuşu ile bu sorunun cevabı yanıtlanmıştır. • tRNA, mRNA’daki kodonlar ile doğru amino asidler arasındaki adaptör moleküldür. t-RNA Yapısı • 75-90 nükleotidi içeren küçük yapılardır. • Yapılarında adenin, guanin, sitozin ve urasil dışında psödouridin, dihidrourasil, timin gibi bazlar da bulunur. Bu yapılar post transkripsiyonel modifikasyonlar sonucu oluşur. • Baz eşlenmelerine bağlı olarak t-RNA ikincil yapı oluşturur. • Modifiye bazlar içeren halkasal bölgelerde baz eşleşmesi bulunmaz. • t-RNA’nın kollarından ikisi adaptör fonksiyonu görür. • Bir tanesi amino asid koludur. Bu kolun 3’ncundaki • • • adenosin kalıntısı spesifik bir amino asid ile esterleşir. Diğeri antikodon koludur. 3.kol D koludur ve dihidroüridin içerir. 4.kol T ψ C halinden ribotimizin psodouridin içerir. Protein Sentez Basamakları • • • • Amino asid aktivasyonu ve tRNA’ya bağlanması Polipeptid zincir sentezinin başlaması Polipeptid zincir sentezinin uzaması Polipeptid zincir sentezinin sonlanması ve polipeptidin ribozomdan ayrılması • Katlanma ve posttranslasyonel işlemler Amino Asid Aktivasyonu • Protein sentezinin ilk fazı sürecinde sitozolde 20 farklı aminoasid tRNA sentetaz enzimleri ile spesifik tRNA’larına bağlanır. • t-RNA’larda antikodonun 3.bazı esnek (wobble) olduğu için 32 farklı t-RNA olduğu düşünülmektedir. • Buna karşılık her amino asid için spesifik bir aminoaçil tRNA sentetaz vardır. Reaksiyon iki safhada oluşur. • 1.safhada amino asidlerin aktif şekli olan amino açil adenilat’lar oluşur. • İkinci safhada ise aktifleşmiş amino asid t-RNA’ya aktarılır. Peptid Zincirinin Sentezinin Başlaması • Prokaryotik hücrelerde polipeptid sentezinin başlaması için: – 30 S ribozomal alt ünite – mRNA molekülü – f met-tRNA – Başlatma faktörleri (IF1, IF2, IF3) – GTP – 50 S ribozomal alt ünite Başlama kompleksinin oluşumu 3 kademede meydana gelir: 1. Kademe • 30 S ribozomal alt ünite iki başlama faktörü bağlar (IF1, IF3) • mRNA 30 S alt üniteye bağlanır. • Başlatıcı kodon AUG’nin önünde bulunan altı bazdan oluşan (AGGAGG) “Shine-Dalgarno” dizisi küçük alt birime bağlanmada rol oynar. • Bakteriyel ribozomlar üç bölge içerir. A P (amino açil) (Peptidil) E bölgeleri (Exit) • Başlatıcı kodon AUG P bölgesinde pozisyon alır. 2.Kademe • Bu kademede 1.kademede oluşmuş olan yapıya GTP bağlı IF-2 ve f-met-tRNA ilave olur. Bu RNA’nın anti kodonu mRNA’nın başlatıcı kodunu ile eş oluşturur. 3.Kademe • Bu kademede oluşan büyük kompleks 50 S ribozomal alt ünite ile birleşir. IF2’ye bağlı GTP hidroliz olur. Üç başlama faktörü ribozomdan ayrılır. • Sonuçta 70 S ribozomdan oluşan başlama kompleksi meydana gelir. Protein Sentezinin Uzama Safhası • Uzama safhası – Başlama kompleksi – Aminoaçil tRNA’lar – Uzama faktörleri (EF-Tu, EF-Ts, EF-G) – GTP içerir. Uzama safhasının 1.kademesinde • Uygun aminoaçil tRNA GTP bağlı EF-Tu kompleksine bağlanır. • Oluşan amino açil-tRNA EF-Tu. GTP kompleksi 70 S başlama kompleksinin A bölgesine bağlanır. • GTP hidroliz olur ve EF-Tu. GDP kompleksi 70 S ribozomdan ayrılır. • EF-Tu GTP kompleksi yeniden oluşturulur. Uzama safhasının 2. Kademesinde; • Ribozomun P ve A bölgelerindeki iki amino asid arasında bir peptid bağı oluşturulur. • Bu reaksiyon peptidil transferaz enzimi ile gerçekleştirilir ve sonuçta A bölgesinde dipeptidiltRNA meydana gelir. Uzama safhasının 3.kademesi Translokasyondur. • Ribozom mRNA’nın 3’ yönünde bir kodon hareket eder. • Bu hareket ile dipeptidil tRNA A bölgesinden P bölgesine kayar. • • • • Deaçil tRNA’da P’den E bölgesine kayar. Bu hareket EF-G (translokaz) gerektirir. Enerji GTP hidrolizinden sağlanır. Ribozom artık 3.aminoasid için hazırdır. Protein Sentezinin Sonlanması • Uzama safhası mRNA üzerinde üç sonlandırıcı kodon (UAA, UAG, UGA)’dan birinin mevcudiyeti ile sonlanır. • Sonlanma kodonunu takiben sonlandırma faktörleri (RF1,RF2,RF3) devreye girer. • Terminal peptidil tRNA bağı hidroliz edilir.