DERS 5 TRANSLASYON (İndirme : 128)
advertisement
")
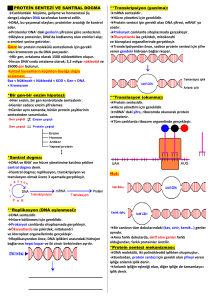
GENETİK ŞİFRE TRANSLASYON VE PROTEİNLER 1 DNA’nın anlamlı zinciri DNA’nın anlamsız zinciri Anlamsız zincirin transkripsiyonu mRNA’nın translasyonu 2 Genetik Şifrenin Özellikleri • 1- Genetik şifre, harfler halinde gösterilen mRNA moleküllerini oluşturan ribonükleotit bazları kullanılarak, doğrusal olarak yazılır. Bu doğrusal yazılım DNA’daki eşlenik bazlardan elde edilir. • 2- mRNA’daki her kelime 3 ribonükleotit harfinden oluşur. Kodon denilen her biri 3 nükleotitlik grup, bir aminoasiti belirler. Bu nedenle şifre üçlüdür (triplet). • 3- Şifre özgündür. Yani her üçlü (kodon) bir aminoasit belirler. • 4- Şifre dejeneredir. Belirli bir aminoasit, birden fazla üçlü kodon tarafından belirlenir. 20 aminoasitin 18’inin birden fazla kodonu bulunur. • 5- Şifrede “başla” ve “dur” sinyalleri bulunur. Bazı üçlüler translasyonu başlatmak, bazıları ise durdurmak için gereklidir. 3 • 6- Şifre içinde noktalama işaretleri (“virgüller”) yoktur. mRNA’nın translasyonu başladığında kodonlar, aralarında bir boşluk ve duraksama olmadan birbirinin ardı sıra okunur. • 7- Şifre üst üste çakışmaz. Translasyon başladıktan sonra, mRNA’nın belirli bir yerindeki herhangi bir ribonükleotit, sadece bir üçlünün parçasıdır. • 8- Şifre hemen hemen evrenseldir. Birkaç küçük istisna dışında, bütün virüsler, prokaryotlar, arkealar ve ökaryotlar aynı şifre sözlüğünü kullanırlar. 4 • mRNA keşfedilmeden önce, DNA’nın doğrudan kendisinin protein sentezini şifrelediği düşünülmekteydi. • 1961 yılında François Jacob ve Jacques Monad mRNA’yı bulmuştur. • 1960 yılında Sidney Brenner 20 aminoasiti kodlayacak şifrenin en az üçlü yapıda olması gerektiğini ileri sürmüştür. Çünkü 4 nükleotitin oluşturduğu şifrede nükleotitler, ikili şifre oluştursaydı, 16 (42) şifre kelimesi olacaktı. Bu 20 aminoasiti şifrelemek için yeterli değildir. • Üçlü şifre yapısı 64 (43) kelime belirleyebilir. 5 Aminoasitlere özgü üçlü özgül dizilerin saptanması ile 2 sonuç ortaya çıkmıştır; 1- Genetik şifre dejerenedir. Bir aminoasit birden fazla üçlü kodon tarafından belirlenebilir. 2- Şifre çok kesindir. Bir üçlü kodon yanlız bir aminoasiti belirler. Şifre sözlüğü 64 çeşit üçlüden oluşur; 61 üçlü kodon aminoasit belirler 3 adet üçlü kodon dur sinyalidir ve hiçbir aminoasit belirlemez. 6 Genetik şifreleme sözlüğü 7 Dejenere Şifre ve Wobble Hipotezi •Aminoasitlerin hemen hepsi 2,3 yada 4 farklı kodon tarafından belirlenir. •Serin,Arjinin, ve Lösin altışar kodon tarafından tanımlanır. •Sadece Triptofan ve Metionin bir kodona sahiptir. •Aynı aminoasiti tanımlayan kodonların ilk iki harfi aynıdır, yalnız üçüncü harfi farklıdır. 1966 yılında Crick, üçüncü pozisyondaki dejenerasyonu gözlemlemiş ve wobble hipotezini önermiştir. – Wobble Hipotezi; • 5’ 3’ yönünde ilk iki nükleotit üçüncüye göre daha kritiktir. • Üçüncü pozisyondaki nükleotit için kodon-antikodon arasında hidrojen bağının kurulmasında esneklik vardır ve baz eşleşmesi kuralına sıkıca uyma zorunluluğu yoktur. Bu hipoteze göre aminoasit belirleyen 61 kodon için en az 30 değişik tRNA gereklidir. Bu translasyonu tehlikeye sokmamak ve enerji tasarrufu açısından da önemlidir. Son tahminlere göre bakterilerde 30-40, hayvan ve bitkilerde 50 kadar tRNA çeşiti bulunmaktadır. 8 Antikodon-Kodon baz eşleşmesinin kuralları İnozin 9 Protein Sentezi Başlama, Sonlanma ve Baskılama • Protein sentezinin başlaması son derece özgündür. Bakterilerde, Metiyonin’in modifiye şekli N-formil metiyonin (fmet) bütün polipeptitlerin yapısına giren ilk aminosittir ve 5’- AUG -3’ kodonu ile şifrelenir. Başlama kodonu olarak da adlandırılır. DNA anlamlı (sense) iplikte 5’- ATG -3’ dir. Protein sentezi tamamlandıktan sonra formil grubu uzaklaştırılır. Ökaryotlarda metiyonin formillenmemiştir. • Polipeptit sentezi için sonlanma kodonları 5’- UAA -3’, 5’- UAG -3’ veya 5’- UGA -3’ dır. Hiçbir a.asiti kodlamaz ve polipeptit sentezi için sonlanma sinyalleridir. Sonlanma kodonları olarak işlev görür. DNA da anlamlı (sense) iplikte 5’- TAA -3’, 5’- TAG -3’ ve 5’- TGA -3’ dizisi ile U hariç aynıdır. Mutasyonla bu kodonlar oluşursa, prematüre (olgunlaşmamış) protein meydana gelir. Böyle mutasyonlar da anlamsız (nonsense) mutasyon olarak adlandırılır. 10 Genetik Şifre Hemen Hemen EVRENSELDİR • mRNA yapısı ve translasyonun işleyişi virüsler, bakteriler, arkealar ve ökaryotlarda büyük oranda benzerdir. • Birçok ökaryotik genin bakterilere aktarılabileceği ve burada transkripsiyon ve translasyonun yapılabileceği, rekombinant DNA teknolojisi sayesinde gösterilmiştir. • Bazı farklılıklar mitokondride ve bazı canlılarda saptanmıştır. Örn. Sonlanma kodonu UGA, insan ve maya mitokondrisinde triptofanı belirler. AGA ökaryotlarda arjinin kodonu iken, mitokondride dur kodonu olarak işlev görür. 11 Evrensel genetik şifrenin istisnaları 12 • mRNA’nın translasyonu, aminoasitlerin polipeptit zincirlerine biyolojik polimerizasyonudur. • Ribozoma bağlanan mRNA’da, belirli bir aminoasite ait özgül bir üçlü kodon bulunur. • Özgül bir tRNA molekülünün nükleotit dizileri arasında ise, kodon ile baz eşleşmesi yapabilen ve antikodon olarak adlandırılan, kodono komplementer üçlü ribonükleotit dizisi bulunur. • tRNA, mRNA’daki özel üçlü kodonlar ile doğru aminoasitler arasındaki adaptör moleküllerdir. 13 Protein sentezine genel bakış • Protein sentezi prokaryotlarda ve ökaryotlarda sitoplazmada gerçekleşir. • Protein sentezi hem ökaryotlarda hem prokaryotlarda ribozomlar tarafından gerçekleştirilir. • Sentezlenen protein prokaryotlarda N-formil metionin, ökaryotlarda metionin aminoasiti ile başlar. Ancak sentez sonrası bazı proteinler değişikliğe uğradığı için olgun protein her zaman metionin ile başlamaz. • Sentez sonrası posttranslasyonal modifikasyona (sentez sonrası değişiklik) uğrarlar. Örn. N-terminalinden bir kısım a.asit dizisinin kesilmesi gibi. • Prokaryotlarda çoğu protein polisistronik mRNA’dan (birden fazla polipeptit zincirini kodlar), ökaryotlarda monosistronik mRNA’dan sentezlenir. Dolayısıyla prokaryotik mRNA’da birden fazla başlama 14 kodonu (AUG) bulunur. • Genetik dizide hem başlama, hemde polipeptit zincirinde bulunan diğer metioninler için kodon AUG dir. Başlama kodonu ve diğer metioninleri ayırdetmek için prokaryotlar başlama kodonunun yaklaşık 5-10 nükleotit gerisinde bulunan spesifik dizi ShineDalgarno dizisi (UAAGGAGG) içerir. Ribozomal 16S rRNA Shine Dalgarno dizisi ile komplementerlik gösterir. Buda ribozomun doğru bir şekilde mRNA ile birleşmesini yönlendirir. 5‘- UAAGGAGG (5-10 baz) AUG mRNA 3'--AUUCCUCC................................. 16S rRNA (ribozom küçük alt birimde) 15 • Ökaryotlarda başlama kodonunun tanınması tam olarak aydınlatılamamıştır. • Ökaryotik 5’cap yapısının başlama bölgesindeki AUG kodonunun tanınmasında etkili olduğu düşünülmektedir. • Ökaryotlarda Marlyn Kozak tarafından bulunan dizinin de başlama bölgesinin tanınmasının etkinliğini artırabileceği düşünülmektedir. 5'--ACCAUGG - mRNA 16 • Nükleotit sırası ve aminoasit sırası arasındaki ilişki Genetik kod olarak adlandırılır. • DNA’daki nükleotit dizisinin polipeptitteki a.asit dizisine taşıyan mRNA’daki 3 lü nükleotit dizileri kodon olarak adlandırılır. Her bir kodon 1 aminoasitin şifresini taşır. • Bazı a.asitler birden fazla kodona sahiptir. • Yine her bir a.asit için kodondaki 3 lü diziye komplementer olan antikodona sahip ve özgül olarak tekbir a.asit taşıyan bir tRNA vardır. • Her zaman peptit Metionin ile başlar ve kodonu 5’- AUG -3’ dir. Başlama kodonu olarak adlandırılır. DNA anlamlı iplikte 5’- ATG -3’ dir. • Polipeptit sentezi için sonlanma kodonları 5’- UAA -3’, 5’- UAG -3’ veya 5’- UGA -3’ dır. Hiçbir a.asiti kodlamaz ve polipeptit sentezi için sonlanma sinyalleridir. DNA’da anlamlı (sense) iplikte 5’- TAA -3’, 5’- TAG -3’ ve 5’- TGA -3’ dizisi ile U hariç aynıdır. • DNA’daki ATG ile başlayıp TAA, TAG ve TGA biten ve genellikle bir polipeptidi kodlayan bölge open reading frame (ORF) = açık okuma çerçevesi olarak adlandırılır 17 18 Protein sentezi 4 aşamada olur • Aminoasitin özgül tRNA’sına bağlanması; A. asit yüklemesi • Başlama • Uzama • Sonlanma 19 Ribozomlar Bakteri, Ökaryot ve Prokaryot Ribozomlarının karşılaştırması Büyük Altünit rRNAs Bakteri (70S) Ökaryot (80S) Mitokondri (55S) 50S 60S 39S 23S (2904 baz) 28S (4700 baz) 16S (1560 baz) 5S (120 baz) 5S (120 baz) 5.8S (160 baz) Proteins 33 ~49 48 Small Subunit 30S 40S 28S rRNA 16S (1542 baz) 18S (1900 baz) 12S (950 baz) Proteins 20 ~33 29 20 • Olgun mRNA 3 ana kısımdan oluşur. – 5’ ucundan, başlama kodonuna kadar olan kısım, 5’ çevrilmeyen Lider Bölge yada kısaca Lider bulunur. – Ortada kalan ikinci kısım okuma çerçevesi olarak adlandırılır. Başlama kodonu ile başlayıp (AUG), üç stop kodonundan birisine (UAA,UAG veya UGA) kadar olan kısımdır. – Son kısım stop kodonlarından sonra 3’ ucuna kadar olan kısmı kapsar ve 3’ çevrilmeyen trailer bölge yada kısaca Trailer olarak adlandırılır. • mRNA uzunluklarındaki farklılık bu üç bölgenin her mRNA çeşitinde farklı olmasıdır. • 5’ AUG UAA 3’ 21 Adenin (A), Guanin (G), Sitozin (C), Urasil (U), İnosin (I) 22 1. Aminoasitin özgül tRNA’sına bağlanması; A. asit yüklemesi • 3’ ucuna aminoasit bağlanmış tRNA, aminoaçil-tRNA olarak adlandırılır. Bu işlemi gerçekleştiren enzim aminoaçil-tRNA sentetaz enzimidir ve ATP’ye ihtiyaç duyar • Bir amino asit tRNA nın antikodonu tarafından belirlenir. Her hücre 20 farklı a.asit için 20 farklı aminoaçil-tRNA sentetaz enzimi içerir. 23 24 2. Başlama (Initiation) • Protein sentezi her zaman metionin ile başlar. Bu nedenle başlangıçta aminoasit taşıyan aminoaçil-tRNA, Met-tRNA’dır. Bakterilerde başlangıç aminoasit formil grubu takılmış olan formilmetionin’dir (fMet). Bu ökaryotlarda bulunmaz. • Önce mRNA’ya uygun bölgeden ribozomun küçük alt ünitesi bağlanır. Bu bağlanmayı yönlendiren küçük alt birimde bulunan 16S rRNA’daki dizi ile mRNA’nın başlama kodonundan (AUG) önceki bölgede bulunan Shine Dalgarno dizisi arasındaki eşleşmedir. İlk aminoasite tRNA bağlanması bunu izler. Bu yapı başlama kompleksi olarak adlandırılır. • Daha sonra bu komplekse büyük alt birimde bağlanması ile başlama aşaması tamamlanmış olur. • Bu aşamada Başlama faktörleri olarak adlandırılan proteinler (IF1,IF2,IF3 25 ve GTP kullanılır. 26 27 28 3. Uzama (Elongation) • Bir ribozom iki büyük tRNA bağlanma bölgesi içerir: A bölgesi ve P bölgesi. İlk a.asiti taşıyan tRNA’nın bağlandığı bölge P bölgesi, ikinci a.asiti taşıyan tRNA’nın yerleştiği bölge A bölgesi adını alır. • P bölgesindeki a.asit ribozom büyük altünitede bulunan Peptidil transferaz enzimi aracılığı ile A bölgesindeki a.asite peptit bağı ile bağlanır. P bölgesindeki tRNA uzaklaşırken, A bölgesindeki tRNA, P bölgesine geçer. Bir sonraki kodona uygun antikodona sahip aminoaçil-tRNA, A bölgesine yerleşir ve bu şekilde zincir uzar. • Bu işlemler sırasında Uzama faktörleri (EF-Tu ve EF-G) adını alan proteinler ve GTP enerjisi kullanılır. 29 30 31 32 4. Sonlanma (Termination) • Protein sentezi, sonlanma kodonlarından (UAA, UAG ve UGA) birisine geldiğinde sonlanır. Bu kodonlar herhangi bir a.asiti kodlamaz. Sonlanma işlemi Sonlanma faktörleri (RF1, RF2, RF3) denilen özel proteinlerce sağlanır. • Bu aşamada polipeptidi taşıyan son tRNA’dan polipeptid ayrılırken, ribozomal alt birimlerde birbirinden ayrılır ve protein sentezi sonlanır. • Ribozomlar tekrar birleşerek yeni bir protein sentezine katılabilir. • Bir mRNA’ya birden fazla ribozom bağlanarak protein sentezini gerçekleştirebilir, bu yapılara polizom adı verilir. 33 34 Ökaryotlarda Translasyon Daha Karmaşıktır 1-Ökaryotlarda daha büyük ribozomlarda gerçekleşir. 2-RNA ve Protein bileşenleri prokaryotlara göre daha karmaşıktır. 3-Ökaryotik mRNA lar prokaryotlara nazaran daha uzun ömürlüdür. Saatlerce protein sentezini devam ettirebilirler. 4-Ökaryotlardaki 5’-cap yapısı mRNA nın daha etkin biçimde translasyonunu sağlar. 5-Ökaryotlarda başlama kodonu AUG çevresinde, kısa bir tanıma dizisi olan 5’-ACCAUGG-3’ bulunur ve Kozak dizisi olarak adlandırılır. Prokaryotlardaki Shine Dalgarno dizisine benzer bir işlev görür. Her iki dizide ribozomal küçük alt birime bağlanmayı kolaylaştırır. 6-Ökaryotlarda formilmetionin yerine protein sentezi Metionin ile başlar. 7-Başlama uzama ve sonlanma faktörleri prokaryotlarla homologtur.Ancak bazı durumlarda dah fazla protein faktöre ihtiyaç gösterir. 8-Ökaryotik ribozomların çoğu endoplazmik retikulum zarına bağlıdır. 35 • Gen hipotezi için önceleri, – Bir-gen : bir-enzim ifadesi kullanılmıştır. Daha sonra – Bir-gen : bir-protein ifadesi kullanılmıştır. Son olarakta ise; – Bir-gen : bir-polipeptit ifadesi kullanılmasının daha doğru bir tanım olduğu anlaşılmıştır. • Protein ve polipeptitte aminoasitlerde oluşur. Aralarındaki fark birarada bulunuş şekilleri ve işlevsel farklılıklarıdır. Polipeptitler protein öncülleridir. • İlk sentezlenen yapı polipeptit zinciridir ve proteinlerin birincil yapısını oluşturur ve düz dallanma göstermeyen bir yapıdır. • Son aldığı konformasyon işlevsel proteini oluşturur. İşlev kazanabilmesi için üç boyutlu yapıyı alması gerekir. • Protein sekonder yapı, α-sarmal (α -heliks) ve β-pileli tabaka olarak ikiye ayrılır. • Üçboyutlu yapı ise proteinin uzaydaki üç boyutlu yapısnı ifade eder. • Kuarternet yapı birkaç polipeptitin zinciri içeren proteinler için geçerlidir ve zincirlerin birbire göre aldıkları konformasyonu gösterir. 36 Posttranslasyonel Modifikasyonlar 1-N-ucu ve C-ucundaki aminoasitler çoğunlukla uzaklaştırılır yada değişime uğrar. Ökaryotlarda N-ucu aminoasiti çoğu kez asetillenir, Prokaryotlarda formil metiyonin enzimatik olarak uzaklaştırlır. 2-Bazen bir aminoasit tek başına değişeme uğrar. Örn. Tirozin gibi aminoasitlerin hidroksil gruplarına fosfatlar takılabilir. Bazı aminoasitlerede metil grubu takılabilir. 3-Bazen karbohidrat yan zinciri takılabilir. Glikoproteinler bu şekilde oluşturulur. 4-Polipeptit zincirlerinke kırpılma olabilir. Örn uzun bir polipeptit zinciri olara sentezlenen insülin kesilerek 51 aminoasitlik son şeklini alır. 5-Sinyal dizileri polipeptitten uzaklaştırılır. N-ucundaki proteinin işlev göreceği yere yönlendirilmesinde rol oynayan 30 aminoasite kadar olan bölge sinyal dizisi olarak adlandırılır ve protein hedeflemesinde (targeting) görevlidir. Protein hedefe ulaştıktan sonra sinyal dizi enzimatik olarak uzaklaştırılır. 6-Polipeptit zincirleri çoğunlukla metallerle kompleks yapmış olarak bulunur. Hemoglobinde 4 demir atomu ve 4 polipeptit zinciri bulunur. • Proteinlerin katlanmalarını şaperonlar adı verilen bir protein ailesi yönlendirir. 37 MUTASYONLAR DNA ONARIMI YER DEĞİŞTİREBİLEN ELEMENTLER 38 • Hergün havadaki, sudaki ve topraktaki binlerce kimyasala maruz kalıyoruz. Bunların birçoğu zaralı. • Bir kısmı ölümcül etkiye sahip. Hastalıklara, doğum hasarlarına ve kansere neden olur. Bazıları da DNA da değişiklikler yani mutasyonlar meydana getirir. • DNA da ne tip mutasyonlar meydana gelir? • Bu tip değişikliklere neden olan kimyasalları nasıl tespit edebiliriz? • DNA mutasyonlarının hayatımızdaki etkileri nelerdir? 39 • Gerçekte mutasyonlara uğrama, genetik maddenin sahip olması gereken özelliklerden birisidir. • Bu değişiklikler bir bazın ilavesi veya eksilmesi şeklinde olabileceği gibi, çok sayıda bazın eklenmesi veya çıkarılması şeklinde de olabilir. • Mutasyonlar genetik çalışmalarda kullanılan önemli araçlardan bir tanesidir. Bazı genlerde meydana getirilen mutasyonlar o genin ifadesi hakkında bilgi edinilmesini sağlar. • Mutasyonlar her zaman olmasa da genellikle spontandır (kendiliğinden). 40 Mutasyonlar birkaç şekilde sınıflandırılabilir – 1. Kromozom Mutasyonları • Kromozom yapısındaki değişiklikler – 2. Genom mutasyonları • Kromozom sayısındaki değişiklikler – 3. Tek-gen mutasyonları • Göreceli olarak küçük bir gen parçasındaki DNA da meydana gelir 41 GEN MUTASYONLARI 1- Spontan mutasyonlar: Doğal olarak meydana gelir. Örn. DNA replikasyonu sırasındaki hatalar. Uyarılmış mutasyonlar: kimyasallar, radyasyon, UV gibi etkenlerin neden olduğu mutasyonlardır. 2-Gametik Mutasyonlar: gamet hücrelerinde meydana gelen mutasyonlardır. Bu mutasyonlar yeni nesillere aktarıldıklarından çok daha önemlidir. Yeni neslin tüm hücrelerinde ifade potansiyeli bulunur. Otozomal dominant mutasyonlar ilk nesilde fenotipik olarak görülecektir. Dişilerin gametlerinde oluşan X’e bağlı resesif (çekinik) mutasyonlar, etkilenmiş X kromozomunu alan hemizigot erkeklerde ifade edilebilir. Somatik mutasyonlar: somatik hücrelerde meydana gelen mutasyonlardır. Bu hücrelerde çekinik aleller oluşturan mutasyonlar genelde önemli değildir. Bu mutasyon dominant (baskın) alel tarafından baskılanır. Somatik mutasyonlar dominant yada X kromozomuna bağlı ise hemen ifade edildikleri için daha büyük bir etkiye sahiptir. Eğer gelişimin erken dönemlerinde meydana gelirse, farklılaşmamış hücrelerdeki mutasyonlar doku ve organlara farklılaştıklarında daha da önemli olacaktır. Yetişkin dokularda oluşan mutasyonlar genellikle mutasyon geçirmemiş milyarlarca hücrenin etkisi ile maskelenecektir. . Erkek ve dişilerin herhangi birinin gametlerinde olusan Otozomal resesif mutasyonlar, heterozigotluk nedeni ile o alel populasyonda yayılana kadar nesiller boyunca dikkat çekmeden aktarılabilir. Şansa bağlı olarak bu iki mutant alel bir araya gelirse, yani homozigot olursa fenotipik olarak bireyleri etkiler 42 İnsanda zararlı mutasyonlar • Mutasyonların çoğunun kodlanmayan geniş genom kısmında yada intron bölgelerinde yer alması olasıdır. Bu bölgelerde meydana gelen mutasyonlar nötral mutasyonlardır. • Zararlı mutasyon sıklığı her nesil başına her birey için en azında 1.6 zararlı genetik değişikliktir ki bu oldukça yüksek bir rakamdır. Mutasyonlar birçok şekilde görülür ve farklı yollarla oluşur • Gen kimyasal bilgiyi temsil eden nükleotit çiftlerinin doğrusal biri dizisidir. • Genetik şifre üçlü nükleotitlerden oluşur. • Üçlü her nükleotit, ilgili proteindeki bir aminoasiti belirler. • Bir harf değişikliği cümlenin anlamını değiştirebilir. – THE CAT SAW THE DOG – THE CAT SAW THE HOG – THE BAT SAW THE DOG • Bunlar yanlış anlam oluşmasına neden olur. Bu tip anlam değişikliği baz yer değiştirmeleri veya nokta mutasyonları olarak adlandırılan mutasyonlara analogdur. Mutasyon bilginin anlamını çeşitli anlamsız formlara dönüştürmüştür. 43 • Substitüsyon da; bir primidin, bir primidin ile yada bir pürin, bir pürin ile yer değiştirmişse bir transisyon (geçiş) olmuştur. • Eğer bir pürin ve primidin karşılıklı yer değiştirmiş ise transversiyon (değişim) olmuştur. – THE CAT SAW THE DOG – THE CAT SAW THE HOG – THE BAT SAW THE DOG • İkinci tip değişiklik, gen içinde herhangi bir noktada bir yada daha fazla nükleotidin girmesi (inversiyon) veya çıkarılmasıdır (delesyon). – THE CMA TSA WTH EDO G – THE ATS AWT HED OG M insersiyonu C delesyonu • Bu tip değişiklikler çerçeve kayması (frame shift) mutasyonlar olarak adlandırılır. Çünkü okuma çerçevesi değişmiştir. 44 a.YANLIŞ ANLAMLI MUTASYON (Missense Mutasyon) Bu tip mutasyonlarda kodondaki nükleotit değişikliği, amino asit değişikliğine neden olur. Etkisi kodon ya da nükleotit tipine göre farklı olabilir.Örneğin, polipeptitteki değişiklik benzer kimyasal özelliğe sahip bir amino asit olarak gerçekleşmişse etkisi daha az, kimyasal özelliği farklı bir amino asit yer almışsa etkisi daha fazla olabilir. DNA-----TGT TGG ACA ACC mRNA- UGU UGG Protein—CysTrp b.ANLAMSIZ MUTASYON (Nonsense Mutasyon) Kodondaki nükleotit değişikliği, normal bir kodonun yerine mRNA’dan translasyonun (protein sentezini) sona ermesini sağlayan “dur” kodonlarından (UAA,UAG ve UGA) birinin oluşmasına neden olur. Polipeptit sentezi erken sonlanır.İşlevsiz bir ürün oluşur. DNA------TGTTGA ACA ACT mRNA---UGUUGA Protein---Cys Dur 45 c.SESSİZ MUTASYON (Silent Mutasyon) Çok hücreli organizmalardaki nokta mutasyonlarının tamamına yakını fenotipik yönüyle nötraldir (insanlarda % 93-97). İnsan DNA’sının yaklaşık %90-97 kadarı “hurda DNA”dan oluşur. Geriye kalan şifre konumunda olan kodonlardaki değişikliklerin 1/3 kadarı sessizdir; kodondaki mutasyonal değişiklik amino asit değişikliğine yansımaz. Çünkü DNA’dan mRNA’ya yansıyan değişiklik, yine aynı amino asiti kodlayan diğer bir kodona dönüşmüştür. DNA------TGTTGC ACA ACG mRNA--- UGUUGC Protein-- CysCys 46 Mutasyona neden olan etkenler Kendiliğinden (Spontan) mutasyonlar – – – – – DNA replikasyonundaki hatalar Tautomerik değişimler(nokta mutasyonu) Bazların Deaminasyonu(nokta mutasyonu) Bazların Depürinasyonu(nokta mutasyonu) Kendiliğinden Metilasyon Mutajenler • Kimyasal Mutajenler • Baz analogları (5-Brom urasil) • Bazların yapısnı değiştiren mutajenler (nitröz asit) • İnterkalasyon yapan ajanlar (akridin boyalar) • Fiziksel mutajenler • İyonize olmayan radyasyon=UV radyasyon (çerçeve kayması) • İyonize radyasyon (X ve gama ışınları gibi) 47