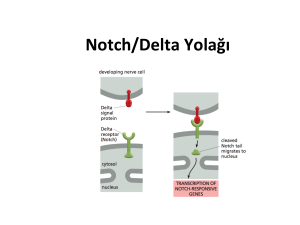
gel***mb*yoloj*s
advertisement

GELİŞİM BİYOLOJİSİ Yard. Doç. Dr. Bedia PALABIYIK Bir hayvan veya bitki yaşamına tek bir hücre (döllenmiş yumurta) olarak başlar Çok sayıda farklı hücre içeren karmaşık ve doğru bir örüntüye sahip olmak için tekrar tekrar bölünür Örüntüyü genomu belirler Gelişim Biyolojisi bu bilmecenin nasıl olduğunu çözmeye çalışır Çok Hücreli bir Organizmanın Sahip Olduğu Hücrelerin Farklılığı • Hücreler aynı genetik bilgiyi taşır. • Her hücrede farklı gen kümelerinin anlatımı yapılır (hücre farklılaşması). Bu süreç embriyoda dört temel işlemi denetler: Hücre Çoğalması Hücre Özelleşmesi Hücre Etkileşimi Hücre Hareketi HAYVAN GELİŞİMİNİN EVRENSEL DÜZENEKLERİ • Yaklaşık on milyon hayvan türünde gelişimin temel mekanizmaların pek çoğu aynıdır. • Özelleşmiş hücre türlerini ve vücut bölgeleri arasındaki farklılıkları belirleyen moleküllerle vücudun biçimsel düzeninin oluşmasını sağlayanlar evrimsel açıdan fark edilir düzeyde birbiriyle ilişkili, benzerdirler. Hayvanlarda Temel Anatomik Özellikler Ortaktır • Gelişimi denetleyen genler hayvan türleri arasında benzerdir (ortak bir atadan) • Bir hayvan vücudu, koruyucu bir dış katman oluşturan epiderm hücreleri, sindirilmiş yiyeceklerden besin emilimini gerçekleştiren sindirim kanalı hücreleri, hareketi sağlayan kas hücreleri, hareketleri denetleyen sinir ve duyu hücrelerinden oluşmaktadır. • Devasa yumurta hücresi bölünür daha küçük boyutta çok sayıda hücre oluşturur. Bu hücreler birbirine yapışır ektodermi oluşturur. Bir yerinden içeri kıvrılır ve endodermi oluşturur. Bir grup hücre ektoderm ile endoderm arasına göç ederek mezodermi oluşturur. Deniz kestanesinde gastrulasyon Çok Hücreli Hayvanlar Hücre Etkileşimlerine ve Gen Düzenlenmesine Aracılık Eden Proteinler Yönünden Zengindir • Moleküler örgütlenme için iki farklı protein sınıfı önemlidir. – Özellikle hücre yapışması ve hücre sinyal iletisinde önemli olan zar geçen moleküller (C.elegans’da 2000, mayada yok) – Gen düzenleyici proteinleri kodlayan genler. Bu proteinler DNA’ya bağlanır. C. elegans’da 41, Drosophila’da 84, insanda 131 ve mayada yalnızca 7 üyesi vardır. Düzenleyici DNA Gelişim Programını Tanımlar • Farklı hayvan grupları benzer gen topluluğuna sahipse, nasıl bu kadar farklı hayvan grupları ortaya çıkıyor? • DNA’da gen dışında bulunan düzenleyici bölgeler ilgili genlerin anlatım yapmasını sağlar ya da engeller. • Hücreler embriyo içindeki konumlarını çevrelerini referans alarak çoğalırken, o an içerdikleri proteinlerin etkinliklerine bağlı olarak, yeni gen gruplarını çalıştırıp bir durumdan bir sonrakine geçerler. Embriyo Çalışmaları Hücreler Arasındaki Etkileşimleri Ortaya Çıkarır • Erişkin bir hayvanın oluşum süreci nasıl incelenir? – Tanımlayıcı embriyoloji • Gelişimi hücrenin davranışı bakımından açıklamak için her hücrenin embriyodaki kendi bölünme, dönüşüm ve göçü izlenebilir • Temeli 19. yy da atılmış. Günümüzde hücre soyu iz sürümü teknikleriyle yapılmakta Xenopus embriyosunda hücre soyu izlenmesi. Erken evredeki (32 hücre) üç hücreye enjeksiyonla farklı floresan boyalar verilmiş ve 10 saat bekletildikten sonra kesit alınmış. • Gelişimi, – Deneysel embriyologlar, hücre ve dokuların çok hücreli yapıları oluşturmak için izledikleri etkileşim yollarını, – Gelişim genetikçileri ise, genlerin etkileri • bakımından analiz ederler. İki yaklaşım birbirini tamamlar. Bir Hücrede Gelişimsel Karar Gözle İzlenebilen Değişikliklerin Çok Öncesinde Verilir • Normal gelişmeye bırakılmış bir embriyodaki belirli bir hücrenin kaderinin ne olacağı, yalnızca yakın gözlem yaparak, iz sürücü boyalar ya da diğer hücre işaretleme teknikleri kullanılarak keşfedilebilir. Hücre kararlılığının standart testi Normal Yazgı Kararsız Kararlı Hücreler Vücut Yerleşimlerini Yansıtan Konumsal Değerlerini Belleklerinde Tutar • Hücreler, özgül bir hücreye farklılaşmaya adanmalarından çok öncesinde, bölgesel olarak kararlı hale gelir. • Bir hücrenin konumuna özgü karakteri, hücrenin konumsal değeri olarak adlandırılır ve etkisini daha sonra gelen örüntü oluşum basamaklarında hücrenin davranış şekliyle gösterir. • Tavukta kanat ve bacak gelişimi. Benzer dokulardan oluşuyorlar. Ne oluyor da farklılık meydana geliyor? Hücre farklılaşması • Gelişimin her evresinde embriyodaki bir hücreye, sınırlı seçenekler sunulur. Gelişimi anlamak için seçenekler arasındaki seçimin nasıl denetlendiğini ve bir önceki seçime nasıl bağlı olduğunu bilmek gerekir. 1) Önemli molekül takımlarının hücre bölünmesi sırasında kardeş hücrelere dengesiz olarak paylaştırıldığı asimetrik hücre bölünmesinin bir sonucu olarak, iki kardeş hücre birbirinden farklı özelliklerle doğar. 2) Komşu hücrelerden gelen sinyaller – Yanal engelleme (Notch yolağı) 3) Aynı gelişim potansiyeline sahip bir grup hücrede, grubun dışındaki hücrelerden gelen bir sinyalin, grubun bir ya da daha çok üyesini farklı bir gelişim yolağına itmesidir. -uyarıcı etkileşim Morfogenler Dereceli Etkiye Sahip Uzun Mesafeli Uyaranlardır • Sinyallerin varlığı ve yokluğunun yanında, ne kadar olduğu da önemlidir. – Örneğin, molekül derişiminin yüksek olması durumunda hedef hücreler belirli bir gelişim yolağına yönelirken, orta düzeydeki gelişim bir başka, daha az düzeydeki derişim de daha başka yolağa yönlendirir. • Hücrenin bulunduğu alanın tamamında belirli bir örüntü oluşturan sinyal molekülleri morfogen olarak adlandırılır. Hücre Dışındaki Sinyal Molekül Engelleyicileri Uyarıcı Yanıtı Şekillendirir • Özellikle uzak noktalarda etkin olabilen moleküller için, sinyal oluşumunu ve etkisini sınırlamak önemlidir. Gelişimsel sinyal proteinlerinin pek çoğunun, işlevlerini engelleyen hücre dışı antagonistleri vardır. • Aslında, şaşırtıcı sayıda gelişim kararı birincil sinyal molekülünün kendisinden ziyade engelleyenler tarafından düzenlenir. Hücrenin İçsel Programı Çoğu Kez Gelişiminin Zaman Sürecini Belirler • Her gelişimsel değişiklik için bir uyarıcı sinyalin gerekli olduğunu düşünmek yanlış olur. • Hücrenin karakterini değiştiren mekanizmaların çoğu, çevresel bir uyaran gerektirmeksizin, hücrenin kendi iç donanımındadır. • Yani hücre sabit bir ortamda tutulsa bile gelişimsel program basamaklarına devam edecektir. İlk Örüntüler Küçük Hücre Alanlarında Oluşur ve Ayrıntılar Embriyo Büyüdükçe Ardışık Uyarılarla Oluşturulur • Embriyodaki uzaysal örüntüyü düzenleyen sinyallerin etkinliği genellikle kısa mesafelidir ve basit seçenekleri yönetir. • İlk komutları izleyen hücre çoğalması boyut artışından sorumludur. İki farklı türden hücre oluşur oluşmaz, bunlardan biri, komşu hücrelerin bir alt kümesini üçüncü bir şekilde özelleşmeleri için uyaran bir etmen üretebilir. Buna karşılık üçüncü hücre ise yakındaki diğer iki hücre tipine sinyal vererek dördüncü ve beşinci hücre türünün oluşumuna yol açar. • Gittikçe daha karışık bir örüntü meydana getiren strateji: ardışık uyarım Caenorhaptidis elegans: HER HÜCRENİN KENDİ BAKIŞ AÇISINDAN GELİŞİMİ • Küçük, nispeten basit ve özenle yapılandırılmış • Gelişim anatomisi tanımlanmış ve vücudundaki her hücrenin soyu kesin olarak haritalanabilmiştir • Tüm genom dizisi bilinmektedir ve mutant fenotipler gen işlevlerini saptamak için analiz edilmiştir • Anatomik açıdan basit olmasına karşın, bilgisayar desteği olmadan izlenemeyecek kadar çok sayıda geni (18.000) ve hücresi (1000) vardır. • Göz ve uzuv gibi organları izlemek için uygun değildir. • Bu kurtçuk başlangıç düzeyi için mükemmel bir örnektir. Caenorhabditis elegans Anatomik Açıdan Basit Bir Organizmadır • • • • • • • 1000 somatik hücre ve 1000-2000 germ hücresi Anterior uçta ağız ve beyin, posterior uçta anus (bilateral simetri ) Vücut yapısı, sinir, kas, bağırsak ve deriden oluşmuştur. Dış vücut duvarı: koruyucu epidermis ve kas katmanı Endodermal hücrelerden oluşan tüp bağırsağı oluşturur. Gonadları meydana getiren ikinci tüp, bağırsakla vücut duvarı arasındadır. C. elegans iki eşeylidir: Hermafrodit ve erkek. Hermafrodit sınırlı sayıda sperm üreten dişidir. Kendileşerek ya da çapraz döllenmeyle üreyebilir. Genetik çalışmalar için önemli Gelişmekte Olan Nematoddaki Hücre Yazgıları Neredeyse Mükemmel bir Şekilde Öngörülebilir • Yaşama döllenmiş bir yumurta halinde başlar ve tekrarlı hücre bölünmeleriyle, yumurta kabuğunun içinde 558 hücreli küçük kurtçuk oluşturur. • Deri değiştirerek dört larva evresi geçirir. • Devam eden bölünmelerle büyür ve cinsel olgunluğa erişir ve kendi yumurtalarını üretmeye başlar. • Yumurtadan yumurtaya gelişimsel süreci üç gün • Tek hücreli yumurtadan çok hücreli erişkin oluşana kadar tüm hücre soyları, gelişen hayvan doğrudan izlenerek haritalanmıştır. • Bu durumdan dolayı uygun model olmadığı ileri sürülmüş • Aksine, gelişim her hücrenin iç işleyişine bağlı olduğu kadar, hücre hücre etkileşimlerine de bağlıdır. • Burada, hücre hücre etkileşim örüntüsü son derece tekrarlanabilir özelliktedir Anasal Etkili Gen Ürünleri Yumurtanın Asimetrik Bölünmesini Düzenler • Gelişimin erken dönemindeki farklar nasıl ortaya çıkar? • Germ hattı, döllenmiş yumurtanın asimetrik bölünme dizisi ile oluşur. • Spermin giriş noktası, uzayan yumurtanın ileri dönemdeki posterior kutbunu tanımlar. • Hücre içinde yumurtadaki proteinler birbirleriyle etkileşir ve kendilerini spermin giriş noktasına göre düzenlerler. Bu işlemde yer alan proteinler genellikle, annenin genlerinden sentezlenmiş mRNA ürünlerinin anlatımı sonucunda üretilmiştir. Bu RNA yumurtlama işleminden önce yapıldığından, bu şekilde etki eden genlere anasal etkili genler denir • Nematodlarda “partitioning defective” par genleri, hücre asimetrisini yapılandırır • Par proteinleri, P tanecikleri olarak adlandırılan bir grup ribonükleoproteini yumurtanın posterior kutbuna getirir. Posterior yavruya P tanecikleri aktarılırken, anterior yavruya aktarılmaz. Birkaç hücre bölünmesinde de bu şekilde çalışır. 16 hücreli evrede P tanecikli sadece bir hücre bulunur. Heterokronik Genler Gelişimin Zamanlamasını Denetler • Hücre içinde yer alan düzenleyici molekül kümesi(leri) de düzenlemeye yol açabilir. • Nematod embriyosunda bile hücre bölünme aşamalarının nasıl denetlendiği hakkında bilinenler azdır. • Ancak, larvaların beslenip büyüdüğü ve erişkin olmak için deri değiştirdiği daha geç evreler için hücresel olayların zamanlamasını denetleyen genlerden bazıları tanımlanmıştır. Bu genlerdeki kusurlar heterokronik fenotiplere yol açar. DROSOPHILA • C. elegans’dan yüz kat daha fazla hücre; 14.000 gen • Sineğin vücut yapısı ve gelişimini denetleyen genlerin bir seti oluşturulmuştur. • Drosopila’da vücut örüntüsünü belirleyen genlerin insan dahil yüksek organizasyonlu canlılarda homologları vardır. Hatta, vücut planını, organ ve dokuların örüntüsünü tanımlayan temel yapılar çok benzerdir. • Çoğaltılması ucuz, mutasyon oluşturulması kolay ve üreme döngüleri hızlı olduğundan genetik çalışmalar için idealdir. Ayrıca, bir karakter- bir gen olduğu için buradaki bir mutasyon doğrudan fenotipte izlenebilir. Sineğin Vücudu Bir Dizi Bölütlenmiş (segmentlenmiş) Birimden Oluşmaktadır Drosopila Gelişimine Ortak Sitoplazmalı Olarak Başlar • Yumurta boyutu 0,5x0,15 mm. • (Dokuz bölünme) ortak sitoplazmalı blastoderm tabakası • (Dört tur çekirdek bölünmesi) yumurta yüzeyinden içeriye doğru çekirdekleri çevreleyecek plazma zarı gelişir ve (6000 hücreli) hücresel blastoderm • Yumurtanın posterior ucuna yerleşen yaklaşık 15 çekirdek birkaç döngü önceden hücrelere ayrışır. Bu kutup hücreleri sperm ve yumurtalara dönüşecek germ hattı öncülleridir. • Hücresel blastoderm evresine kadar, yumurtada biriken anasal mRNA ve protein stokları gelişimi yönetir. • Hücreleşmenin tamamlanmasından kısa bir süre önce hücre katmanının bir kısmı içeriye sokulmaya başlar (gastrulasyon) başlar. • Gastrulasyon tamamlanmaya başladıkça embrio yüzeyinde vücudun alt bölmelerini anteroposterior eksen boyunca bölütler (segmentler) halinde işaretleyen girinti ve çıkıntılar görünmeye başlar. Yeme ve büyümeye başlamak için hazır larva meydana gelir. Erken Örüntüleme İçin Gerekli Gen Grupları Gap genleri Çift-kural “pairrule” genleri “segment-polarity” bölüt-kutup genleri • En az altı üyeli • Embriyoyu kabaca bölümlere ayırır ve sınırlarını çizer • Kruppel mutantı • Sekiz üyeli • Birbirini izleyen bir dizi segment kaybolur ve mevcut segmentin yarısı kalır. • Even skipped (eve) ve fushi tarazu (frz) mutantları • On üyeli • Delesyona uğrayan her parçanın yerini, geriye kalan parçanın ayna kopyası alır. • Gooseberry mutantı “Homeotic • Komşu segmentler (bölütler) selector” arasındaki farkı tanımlama homeotik ve korumayı sağlar seçici genler Oositin Çevresiyle Etkileşimi Embriyo Eksenini tanımlar: Yumurta kutuplaşma Genlerinin Rolü • Embriyonun gelecekteki anterior ucunda Bicoid mRNA’sı bulunur. Bicoid protein gradyanı oluştururlar. anterior • Embriyonun gelecekteki posterior ucunda Nanos’a ait mRNA’lar posterior gradyanı oluşturur. posterior • Torso’nun yerel olarak etkinleşmesiyle, gelecekte larvanın baş ve kuyruk uçlarını oluşturacak özelleşmiş uç yapılarının yerleri ile sindirim kanalının temel yapılarını belirler. Etki alanı kısadır. terminal • Dorsoventral ekseni tanımlar. Embriyonun hemen altındaki folikül hücrelerinin ürettiği bir protein, oosit zarında bullunan, Toll reseptörünün yerel etkinliğine yol açar. Burada görevli genler dorsoventral yumurta kutuplaşma genleri • Bicoid, Nanos, Torso ve Toll tarafından sağlanan her kutuplaşma sinyali, etkisini doğrudan veya dolaylı olarak blastoderm çekirdeklerindeki genlerin anlatımını düzenleyerek gösterir. Böcek Dorsaventral Ekseni Omurgalı Ventrodorsal Eksenine Karşılık Gelir • Omurgalılarda, Drosophila’da bulunan Dpp ve sog proteinlerine benzer şekilde çalışan BMP4 ve kordin proteinleri vardır. Üç Sınıf Segmentasyon Geni Anterior Posterior Anasal Örüntüyü Ayrıntılandırır ve Embriyoyu Alt Bölümlere Ayırır • Bu genler embriyonun kendi genomunun ürettiği ilk genlerdir. Zigotik etkili genler. Segmentasyon genlerinin Lokal Anlatımı Sinyal Hiyerarşisiyle Düzenlenir • Her aşamada sırasıyla yeni ayrıntıları tanımlamak için yeterli düzeyde derişim farkları sağlayan yeni sinyaller ortaya çıkar. • Böylece, biyolojik denetim sistemindeki düzensizliklere ve sineğin gelişimi sırasında değişen çevresel koşullara karşın, tümüyle aynı örüntüye sahip sinek embriyoları oluşturulabilir. HOMEOTİK SEÇİCİ GENLER VE ANTEROPOSTERİOR EKSENİN ÖRÜNTÜLENMESİ • Gelişim ilerledikçe vücut daha karmaşık hale gelir. • Her türde ve yapılanmanın her düzeyinde, karmaşık yapıların her zaman az sayıdaki temel olgunun küçük değişikliklerle tekrarlanmasından oluştuğunu görüyoruz. – Kas, fibroblast – Çeşitli dokuların bir araya gelmesinden, diş veya parmak gibi organlar ortaya çıkar. • Bu modüler tekrarlamanın olduğu yerde iki soru: Belli bir grup nesnenin tamamında ortak olan temel yapım mekanizması nedir? Ve bu mekanizma gözlemlenen farklılıkları oluşturmak için nasıl değişkenlik göstermektedir? • Segmentlerin temelinin ortak ve ayrı özellikleri ORGANOGENEZ VE UZUV ÖRÜNTÜLENMESİ • Böcek vücudundaki başlıca uzantılar ve diğer pek çok vücut parçası, evrimsel olarak korunmuş, az sayıdaki temel temanın değişik biçimleri olan alt yapılardan oluşmaktadır. Erişkin Sineğin Vücut Bölümleri İmge Disklerinden Gelişir • Erişkin sinekteki dış yapılar, larvanın her segmentinde bulunan, görünürde farklılaşmamış hücre gruplarının oluşturduğu, imge diskleri “imaginal discs”den gelişmiş yapılardan köken alır. • Diskler sinmiş, buruşuk balon görünümündeki epitel torbacıklardan oluşur ve larvanın epidermis yüzey katmanıyla devamlılık gösterir. Bunlardan her yanda dokuzar çift ve ortada bir adet olmak üzere 19 adet vardır. Uzuvları Oluşturacak Hücreleri Özgül Düzenleyici Genler Tanımlar • İşlem erken dönem örüntülenme mekanizmaları ile başlar. • Moleküler anlamda imge diski-tanıyıcı düzenleyici genlerin açılmasına karşılık gelmektedir. Disklerin çoğunda Distal-less geni açılır. Bu gen anten veya bacak gibi proksimodistal eksenli uzantıları oluşturmak üzere devamlı büyüme için gerekli olan düzenleyici protein genini kodlar. Genin yokluğunda bu tür uzantılar oluşmaz, gen olağan dışı bölgelerde ifade edilirse yanlış yerleşimli uzantılar oluşur. Göz diski için de eye-less geni benzer işlev yapar. Deniz kestanesi Güve larvası Böcek Kanat Diski Bölmelere Ayrılmıştır • İmge diskini oluşturan hücre kümeleri başlangıçtan itibaren, henüz tam gelişmemiş iç örüntüleri taşırlar. Kanat diskinin posterior yarısında bulunan hücreler engrailed segment kutuplaşma genini ifade ederken, anterior yarıda bulunan hücrelerde bu gen ifade edilmez. Kanat Örüntüsünü İyi Bilinen Dört Sinyal Yolağının Bileşimi Oluşturur: Wingless, Hedgehog, Dpp ve Notch • Her bölmenin sınırları boyunca uzanan farklı durumdaki hücreler birbirleriyle karşılaşır ve özelleşmiş hücrelerden oluşan dar bantları oluşturmak için etkileşir. Bu sınır hücreleri daha sonraki büyümeyi sağlamak ve uzantıların ayrıntılı örüntüsünü sağlamak için yeni sinyaller üretir. – Posterior kanat bölmesindeki hücreler Hedgehog sinyal proteinini ifade eder ancak buna yanıt veremez. – Anterior bölmedeki hücreler buna yanıt verebilir. Hedgehog etki alanı kısa olduğundan sınırdaki hücre bantı etkin olur. – Bu sınır hücreleri Dpp sinyalini de başlatır. Sınır hücrelerinden dışarıya doğru difüzyonla yayılarak bir morfogen gradyanı oluşturur. Bu şekilde sonraki örüntüyü denetleyebilir. • Benzer olaylar dorsoventral sınırda da gerçekleşir. Notch yolağının aracılığıyla diğer bir morfogen olan Wingless proteinini üreten bir sınır hücre bandı yaratır. Her Bölmenin Boyutu Hücreleri Arasındaki Etkileşimlerle Düzenlenir • Neden vücudun her bölümü kesin olarak belirlenmiş bir boyuta kadar büyümektedir? • Bölme içindeki hücreler birbirleriyle etkileşerek bölmenin sonlandırılacağı zamanı belirlemekte • Hücre bölünmesini yavaşlatan yada hızlandıran mutasyonlarda, hücre boyutu ve sayısı değişerek organ boyutunun sabit • İnsülin gibi büyüme etmenlerinin aracılık ettiği sinyal yolaklarındaki genetik kusur: az sayıda hücre içeren küçük sinekler, aşırı etkinlik: dev sinekler Erişkin kanatta işaretlenmiş kolonilerin şekli engriled geninin kanattaki anlatımı • Büyüme hızındaki veya başlangıç koşullarındaki yerel bozukluklara karşın bir bölmenin içindeki her küçük örüntünün uygun boyutlara büyümesini sağlayan mekanizma nedir? – Morfogen gradyantları farklı konumdaki hücrelere farklı özyapılar yüklemektedir. • Gelişmekte olan hamam böceğinde farklı ayak parçaları birbirine nakledildiğinde meydana gelen ara doldurucu yeniden oluşumla bacağı olağan yapı ve boyutuna uygun olarak onarır. Omurgalılarda Uzuv Örüntülenmesini de Benzer Mekanizmalar Sağlar • Omurgalılardaki kol ve bacak uzantıları böceklerinkinden çok farklıdır. Ancak, moleküler mekanizmalar oldukça benzerdir. • Tavuğun bacak ve kanatlarından her biri uzuv tomurcuğundan kökenlenir. Drosophila kanat disklerindeki Distal-less, wigless, Notch, engrailed, dpp ve hedgehog genlerinin benzeşikleri bu yapının içinde anlatılır edilir. • Hox genleri de omurgalı uzantılarında rol oynar. Tüm vücudun anterioposterior ekseni boyunca Hox karmaşım genlerine ait farklı genler ardışık anlatımları yapılır. Özgül Gen Düzenleyici Protein Sınıflarının Yerel Anlatımı Hücre Farklılaşmasının Habercisidir • Mekanik duyu kılı dört çeşit hücreden oluşur: mil “shaft”hücresi, yuva “socket” hücresi, nöral kılıf “sheat” hücresi ve nöron. • Erişkin sinekteki kıl hücreleri imge diski epitelinden kökenlenmiştir. Tek bir ana duyu hücresinin ikinci ve üçüncü kuşak torunlarıdır. • İmge diski epitelinde kıl oluşumunu başlatmada achaete ve scute genleri rol oynar. Bu genlerin anlatımı geçicidir ve başlangıçta anlatımı yapılan hücrelerden çok azı ana duyu hücrelerine dönüşür. Diğerleri sıradan epidermis olur. achaete ve scute genleri pronöral genler Yanal Baskılama Ana Duyu Hücresinin Pronöral Hücre Kümelerinden Seçilerek Ayrılmasını Sağlar • Pronöral genlerin anlatım yapıldığı hücreler imge diski epitelinde gruplar halinde bulunur. • Ana duyu hücresi, pronöral genlerin anlatımını kapatmış ana duyu yerine epidermise farklılaşmaya mahkum edilmiş hücrelerle çevrilir. (Notch sinyal ileti yolağı arcılığı) Yanal Baskılama Ana Duyu Hücresi Soylarını Farklı Son Yazgılara İter • Notch yolağına dayanan aynı yanal engelleme mekanizması, duyu kıllarının oluşumunda tekrar tekrar çalışarak yalnızca ana duyu hücresinin komşularını epidermal hücre olmaları için farklı bir yolağa zorlamakla kalmaz; aynı zamanda, ana duyu hücresinin yavrularının, torunlarının ve büyük torunlarının duyu kılına ait farklı bileşenleri oluşturabilmeleri için, farklı genleri anlatmalarını da sağlar. • Numb proteini bölünen hücrenin bir tarafında yerleşim göstermeye başlar ve böylece Numb proteini hücrelerden birine aktarılırken diğerine aktarılmaz. Numb Notch ile etkileşime girerek onun etkinliğini durdurur. Numb bulunduran hücre komşudan gelen sinyale sağır kalırken, kardeş hücreler varlıklarını korur. Asimetrik Bölünmenin Düzlemsel Kutuplaşması Frizzled Reseptörü Yoluyla Sağlanan Sinyal İletimi İle denetlenir • Ana duyu hücresinin geçirdiği ilk bölünmede, Numb anterior hücreye aktarılırken, posterior hücreye aktarılmaz (düzlemsel kutuplaşma). Bu durum sineğe, duyu kıllarının hep birlikte arkaya doğru yatık durmasıyla kendini gösterir. • Ana duyu hücresini başlangıç bölünmesindeki düzlemsel kutuplaşma, Frizzled reseptörüne bağlı olarak denetlenmektedir. Burada hücre içi nakil düzeneği, etkilerini gen anlatımı yerine, aktin hücre iskeleti düzeneğinde göstermesine yol açar. Frizzled’ın aşağı tarafında bulunan Dishevelled proteini, sinyal yolağının gen düzenleyici ve aktin kodlarında ortaktır. Yanal Baskılama ve Asimetrik Bölünme Vücutta Boydan Boya Nöron Oluşumunu Birlikte Düzenler • Hem sineklerin hem de omurgalıların embriyonik merkezi sinir sisteminde nöronlar, achaete ve scute benzeri pronöral genlerin anlatıldığı bölgelerde oluşur. Yeni oluşan nöron öncülleri Delta’yı anlatır ve Notch anlatan yakın komşularının nöral farklılaşmaya adanmasını engeller. • Nörogenez işlemlerinin pek çoğunda asimetrik hücre bölünmesi önemli rol oynar. Apikal yerleşimli bir protein (Bazooka)nin omurgalı merkezi sinir sisteminin gelişiminde etkisi vardır. Nematod embriyosundaki Par proteininin Par 3 genine benzerdir. Notch Siyal İletimi Pek Çok Farklı Dokudaki Farklılaşmış Hücre Tiplerinin İnce Örüntüsünü Düzenler • Notch sinyal iletimi, kasta, sindirim kanalı astarında, salgı sisteminde, trakede, göz ve diğer duyu organlarında da denetleme yapar. • Bu dokuların hepsinde farklı hücre tiplerinin dengeli bir karışımına gereksinim vardır. Notch sinyal iletimi, bunu sağlamada bir araçtır. Bazı Anahtar Düzenleyici Genler Hücre Tipini Tanımlar; Diğerleri Bütün Olarak Bir Organın Oluşumu İçin Gerekli Programı Etkinleştirebilir • Belli bir farklılaşma, Notch aracılığıyla oluşan etkileşimlerden sonra, özgül hücre farklılaşması genlerinin anlatılmasıyla belirlenir. Çok daha az sayıdaki yüksek düzey düzenleyiciler tarafından sağlanır. Bunlar, MyoD ailesinden MyoD, myogenin, Myf5, MRF4 ve omurgalılardaki benzeşikleridir. • Çok karmaşık gibi görülen gelişim genetiğinin en önemli mesajı, beklenmedik düzeyde basittir. Bütün bu hayvan gelişiminin temel özelliklerini, farklı koşul ve bileşimlerle tekrar tekrar kullanılan sınırlı sayıda gen ve mekanizma denetlemekten sorumludur. HÜCRE HAREKETLERİ VE OMURGALI VÜCUDUNUN ŞEKİLLENMESİ • Hayvan vücudundaki hücrelerin çoğu hareketlidir. • Gen anlatımındaki denetimli değişiklikler, farklı hallerde bulunan düzenli hücre sıraları yaratır. • Bu da hücresel yapıtaşlarını yeniden düzenler ve olması gereken yerlerine koyar. Amfibi Embriyosunun Kutuplaşması Yumurta Kutuplaşmasına Bağlıdır • Xenopus yumurtası, büyük bir hücredir. • Açık renkli alt ucu bitkisel (vejetal) kutup, koyu renkli üst ucu hayvansal (animal) kutup. • Vejetal kutupta VegT ve TGF üst ailesinin sinyal iletim proteinlerini kodlayan mRNAların yanı sıra, Wnt sinyal ileti yolağının hazır proteinleri bulunmaktadır. • Vegetal sitoplazmayı alan hücreler, vücudun en iç dokusu olan sindirim kanalını oluşturmaya adanırken, animal sitoplazmayı alan hücreler dış dokuları oluşturur. • Döllenme bir dizi karışık hareketi başlatır. Üç temel eksen oluşur: baştan kuyruğa anterioposterior, sırttan karına dorsoventral, orta hattın sağından veya solundan dışarıya doğru mediolateral. • Yumurtanın animal-vejetal asimetrisi anterşoposterior ekseni • Döllenme hücre içi bir hareketi tetikleyerek dorsoventral asimetri kazandırır. Sperm girişini takiben , yumurta sitoplazmasının aktince zengin dış kabuğu, yumurta merkezine göre döner ve böylece kabuğun animal kutbu hafifçe gelecekteki ventral tarafa kayar. Kabuk dönüş yönü, sperm giriş noktasına göre yönlenir ve hareket, yumurta sitoplazmasındaki mikrotübüllerde meydana gelen yeni bir yapılanmayla bağlantılıdır. Bu Wnt sinyal yolağının aşağı yönde bir bileşeni olan Dishevelled proteininin mikrotübüller aracılığıyla gelecekteki dorsal tarafa doğru taşınmasına yol açar. Yarılma Bir Hücreden Pek Çok Hücre Üretir • Kabuk dönüşü döllenmeden 1 saat sonra tamamlanır. Bunu hızlı ve tekrarlı mitozlar izler. Böylece blastomerler denilen çok sayıda küçük hücreye yarılır. • Bu yolla, yumurtada asimetrik olarak dağılmış belirleyiciler, farklı yazgıları olan farklı hücrelere dağıtılmış olur. • Xenopus’ta İlk hücre bölünmesi yaklaşık 30 dakikadır. • S ve M evreleri doğrudan birbirini izler. • Yarılma geçiren embriyo yumurtanın annede oosit olarak gelişimi sırasında depolanan RNA, protein, zar ve diğer birikmiş içeriğe bağımlıdır. • Yaklaşık 12 yarılma döngüsünden (7 saat) sonra hücrelerin bölünme hızı yavaşlar. • Hücre döngüleri S ve M evrelerinin arasındaki G1 ve G2 evreleriyle olağan hale gelir. • Bundan sonra embriyo genomunun yazılımı başlar. (ara blastula geçişi ) Gastrulasyon İçi Boş Bir Hücre Topunu Bir Sindirim Kanalı İle Üç Katmanlı Bir Yapıya Dönüştürür • Kurbağa embriyosu, sıvıyla dolu iç boşluğu olan ve birbirine sıkıca bağlı hücrelerden oluşan epitel örüntüyle çevrilmiş topa benzer bir yapıya dönüşür. (blastula) • Gastrulasyon, blastulayı merkezinde sindirim kanalının tüpü bulunan, bilateral simetrili ve çok katmanlı bir yapıya dönüştürür. • İçteki endoderm ilkel sindirim kanalını oluşturmak için içeri hareket eden hücrelerden; dıştaki ektoderm dış kısımda kalan hücrelerden ve her ikinin arasında kalan mezoderm, daha gevşek yapıya sahip bağ dokusunu oluşturmak için epitelden ayrılan hücrelerden oluşur. Erişkin omurgalı vücudunun dokuları, bu üç germ katmanından oluşacaktır. Gastrulasyon Hareketleri Tam Olarak Öngörülebilir • Gelecekteki endoderm hücreleri ardışık olarak yuvarlanıp içe doğru kıvrılır, dürülür. Vejetal kutbun yakınındaki hücreler önce içe doğru katlanarak dürülür ve daha sonra, sindirim kanalının en anterior ucunu oluşturacak biçimde yukarı doğru hareket eder. Hayvansal kutba yaklaşan bu öncü endoderm hücreleri, dışta uzanan ektoderme sinyal ileterek, başın anterior ucunu tanımlayacaktır. Sonuçta, endoderm ve ektodermin doğrudan temas ettiği anterior kısımda , bir boşluk (ağız) gelişir. Bu arada, geleceğin mezoderm hücreleri, ekdoderm ve endodermin arasını doldurmak için epitel örüntüden ayrılır, endoderm hücreleriyle birlikte içeriye sokularak, hayvansal kutup yönünde hareket eder. İçeriye ilk kıvrılan hücreler başın bölmelerini, en son kıvrılanlar kuyruğun bölümlerini meydana getirir. • Vücudun dorsoventral eksenini örüntüleyen hareketlerle, anterioposterior hareket birlikte ilerler. Gastrulasyon, kabuk dönüşü ile blastulanın dorsal tarafı olarak işaretlenen bölgede başlar. Burada, hücrelerin içe doğru kıvrılması, blastopor (bitkisel kutbu çevreleyen bir girinti hattı) oluşturmak için hızla uzayan küçük bir girintiyle başlar. Girintileşmenin başladığı yer dorsal blastopor dudağını tanımlar. Mekanik İşlemleri Kimyasal Sinyaller Tetikler • Gastrulasyon herektleri vejetal blastomerlerden yayılan sinyaller atrafından tetiklenir. TGF üst ailesinin birkaç proteini bu hücreler tarafından salgılanır ve bunların yukarısındaki blastomerlere etki eder. Wnt sinyal yolağına ait bileşenlerin, embriyonun dorsal bölgesinde yerel olarak etkinleştirilmesi, diğer sinyallerin etkinliğinde değişikliğe yol açar ve böylece, dorsal blastopor dudağını oluşturan özel hücrelerin gelişimi uyarılır. • Dorsal blastopor dudağı gastrulasyonda yalnızca geometrik açıdan merkezi bir rol oynamakla kalmaz, aynı zamanda güçlü bir denetim kaynağı olarak da rol oynar. Blastoporun dorsal dudağı gastrulasyonun başında kesilip başka bir embriyonun farklı bir yerine aşılanırsa, embriyo hem kendi dorsal dudak alanında hem de aşılanan bölgede gastrulasyonu başlatır. F.6B Siyam ikizi. • Vücudun bu çok önemli ana ekseni düzenleme rolü nedeniyle, dorsal blastopor dudağı Örgütleyici (Speman) olarak bilinir. Etkin Hücre Paketleme Değişimleri Gastrulasyona İtici Güç Sağlar • Gastrulasyon, blastopor bölgesindeki hücrelerin biçim değiştirmesiyle başlar. Bu hücreler amfibilerde şişe hücreleri olarak adlandırılır: Geniş vücutları ve kendilerini epitel yüzeyine tutunduran ince boyunları vardır. Bunlar içeri bükülmede itici bir kuvvet uyguluyor olabilirler. Örgütleyici civarında yakınsayan genleşmeler geçekleşir. • Bu olağan üstü dönüşümü meydana getirmek için birbiri üzerine sürünerek hareket etmeleri gerekmektedir. Hareketlerin aynı hizada gerçekleşmesi, Frizzled/Dishevelled kutuplaşma sinyal ileti yolağına dayanıyor. Hücre Yapışma Molekülleri Örüntüsünün Değişimi Hücreleri Yeni Düzenlemelere Zorlar • Erken dönem amfibi embriyolarında gelişi güzel karıştırılan hücreler, seçici hücre yüzey etkisiyle, aslına uygun olarak kendiliğinden bir araya gelir. • Kaderinler evrimsel olarak birbiriyle ilintili çeşitli Ca2+ bağımlı hücre yapışma proteinlerinden oluşan, büyük bir ailenin üyeleri. • Gastrulasyon, norulasyon ve somit oluşumu sırasında hücrelerin bağlantı örüntülerinde meydana gelen değişikliklerle, çeşitli kaderin moleküllerinin ifade edilmesindeki örüntüler ilişkilidir. Kaderinler yalnızca bir hücreyi diğerine yapıştırmakla kalmaz, aynı zamanda hücre hücre yapışma yerlerinde hücre içi aktin iplikçikleri için bağlanama noktaları sağlar. Gelişmekte olan dokudaki gerilim ve hareket örüntüleri, yapışma örüntülerine göre düzenlenmektedir. Nöral Plaka Nöral Tüpü Oluşturmak Üzere Kıvrılırken, Notokord Uzar • Yeni gastrulasyon geçirmiş bir embriyoda, mezoderm tabakası vücudun sağ ve solunda iki dilime ayrılır. Mezodermin notokord olarak adlandırılan en erken evre özelleşmesidir. Merkezi vücut eksenini tanımlar ve bu ayrımı gerçekleştirir. Notokord hücreleri, gen düzenleyici bir protein olan Brachyury proteininin ifade edilmesiyle tanımlanır. • Notokord hücreleri dorsal blastopor dudağının etrafından geçip embriyonun içine hareket ederken, yakınsayan genleşme yoluyla çarpıcı derecede uzayan bir doku sütunu oluşur. Notokord hücrelerinin vakuolle şişirilmesi, kolonun daha da uzamasını ve embriyonun gerilmesini sağlar. • Bu arada, sinir sistemini oluşturmak için, ektoderm örüntüsünde başka hareketler meydana gelmektedir. Nörülasyon olarak bilinene bu işlemle, ektodermin nöral plaka olarak adlandırılan geniş merkez bölgesi kalınlaşıp, tüp yapısı oluşturacak şekilde kıvrılır ve kendisini kaplayan hücre örüntüsünden kopar. Ektodermden oluşan bu yapı nöral tüp olarak adlandırılır. Nöral tüp gelişimin sonraki evrelerinde, beyin ve omuriliği oluşturur. Gen Anlatımı Salınım Düzeneği Mezodermin Somitlere Bölünmesini Denetler • Yeni oluşan nöral tüpün her iki yanında birer mezoderm dilimi bulunur. Bu dilimler ardışık omurga dizisini, kaburgaları ve kas bölütlerini oluşturmak üzere, somit denilen, yarıklarla birbirinden ayrılmış hücre gruplarına ayrılır. Tavuk embriyosunda somitler kafadan başlayıp kuyrukta sonlanacak şekilde, birbiri ardına oluşur. Somitlerin toplam sayısı 50’den az (kurbağa, kuş) veya 300’den fazla (yılan) olabilmektedir. • Yeni bir somitin posterior bölümünü oluşturmak üzere hücreler bir grup genin anlatımını başlatırken, bir sonraki somitin anterior bölümünü oluşturacak olan hücreler diğer bir gen grubunu anlatmaya başlar. Kaderinleri ifade eden ve etmeyen hücrelerin karşılaştığı yerlerde somitler arasında yarıklar oluşur. • Her yeni oluşan somitteki hücrelerin çoğu, hızla farklılaşarak, ana vücut ekseninin kas bölütlerine karşılık gelen kas bloklarını oluşturur. Artık bu aşamada embriyo kıpırdayabilir. Somit hücrelerinin farklı alt grupları, omurlar ve dermis gibi bağ dokularını oluşturacaktır. Başka bir alt grup somitten ayrılır ve diğer hücrelerin arasından sürünerek geçip, bölütlere ayrılmamış mezoderme göç eder. Bu hücreler, uzuvlarda dahil olmak üzere, vücutta hemen tüm iskelet kası hücrelerini oluşturacaktır. Göç Eden Hücreler Embriyonik dokulara Sıkı Denetim Altında Yayılır • Somitlerden göç eden kas hücresi öncülleri (miyoblastlar) kararlı olmalarına karşın açık bir biçimde farklılaşmamışlardır. Koloniler kurdukları dokularda, görünüşte kendilerinden hemen ayırt edilemeyen hücre sınıflarıyla karışık halde bulunurlar. Ancak, miyoblasta özgü gen düzenleyici proteinleri (MyoD) anlatımını sürdürürler. Farklılaşmada yalnızca miyoblastlar kas hücresine dönüşecektir. • Embriyonik bağ dokusu, miyoblastların içinde göç ettiği bir çatı oluşturur ve dağılımlarına rehberlik eden sinyalleri üretir. • Bu arada, diğer göç eden hücre sınıfları göç için farklı yollar seçer. Nöral tüpün gelecekte epidermisten kopacağı hat boyunca, bir dizi ektodermal hücre epitelden kopar ve bu hücreler tek tek mezodermin içinden göç eder. Bunlar nöral tepecik hücrelerdir ve periferik sinir sisteminin neredeyse tüm nöronlarını ve glial hücrelerini, derinin pigment hücrelerini. Kafatası ve çene kemikleri de dahil başta bulunan çok sayıda bağ dokusunu meydana getirir. • Diğer önemli göç eden hücreler; kan hücrelerinin ve germ hücrelerinin öncüllerini, merkezi sinir sisteminde bulunan çok sayıda nöron gruplarını ve kan damarlarını oluşturan endotel hücreleridir. Bu göç gruplarından her biri farklı yerlerde koloniler oluştururlar. Böyle bir ayırımın sonunda, omurgalı vücudunun pek çok dokusu, embriyonun geniş alana yayılan farklı bölgelerinden oluşmuş farklı karakterdeki hücre karışımlarından oluşur. • Göç eden hücre embriyonik dokular boyunca yol alırken yüzeyinden dışarı uzantılar çıkartır. Bu almaç proteinler, hücrenin hareket etmesini sağlayan iç kısımdaki hücre iskeletine bağlanmıştır. – Fibronektin proteini gibi bazı hücre dışı matris öğeleri, hücrenin ilerlemesine yardımcı olan tutunma bölgeleri sağlar. – Kondroitin sülfat proteoglikan gibi diğer bazı moleküller de hareketi sınırlayarak, göçü engeller. • Yol boyunca yer alan sabit hücrelerin de benzer şekilde, itici veya çekici yüzeyleri olabileceği ve hatta filopod uzantılarıyla göç eden hücrelere dokunarak bu hücrelerin davranışlarını etkilediği düşünülmektedir. Göç Eden Hücrelerin Dağılımı Yönlendirici İzlere Olduğu Kadar Sağkalım Etmenlerine de Bağlıdır • Göç eden hücrelerin nihai dağılımı yalnızca seçtikleri yollara değil, aynı zamanda yolculuk boyunca hayatta kalmalarına ve yolculuk sonunda varılan noktadaki çevre koşullarında gelişebilmelerine bağlıdır. • Germ hücreleri, kan hücresi öncülleri ve nöral tepecikten köken alan pigment hücrelerinin hepsi, sağkalım için en az bir ortak koşulu paylaşır. Göç eden hücrelerin zarında bulunan Kit proteini. Bu proteinleri kodlayan genlerden birinde mutasyon taşıyan bireylerde, pigmentasyon, kan hücresi ve germ hücresi üretimi yetersizdir. FARE • Uterusta geliştiği için çok küçük ve ulaşılmaz olmasına rağmen – Memeli olması – Küçük boyutları ve hızlı üremesi Üzerinde çok çalışılan uygun bir modeldir. İnsandan evrimsel süreçte yalnızca 100 milyon yıl önce ayrılmıştır. Memeli Gelişimi Özelleşmiş Bir Ön Hazırlıkla Başlar • • • • • Anne karnında besin ihtiyacını plasentadan karşıladığı için yumurta, yolk depolamak zorunda değildir. Yumurtanın çapı 80 µm dir (amfibininkinden 2000 kat küçük) Yarılma bölünmeleri somatik hücrelerin bölünmelerinden hızlı değildir ve gen yazılımı iki hücre evresinde başlar. Memeliler amniyotik kese ve plasenta gibi gerçek embriyoyu çevreleyen, koruyan anne ile arasında karşılıklı metabolit alışverişi sağlayan bir dizi karmaşık yapıyı oluşturarak farklı bir embriyotik gelişim başlatırlar. Döllenmenin 3. gününde 16 hücre oluşur. • Hücreler önce gevşek sonra sıkıdır: Morula. Morula içi boş bir küre olan blastosite “blastocyst” dönüşür. Dış hücre katmanları (trofektoderm, “trophectoderm”) embriyo dışı dokuları meydana getirecektir. İç hücre kitlesi boşluğun bir tarafında yerleşir. Gerçek embriyonun tamamını meydana getirir. • Trofektoderm hücreleri uterus duvarıyla temas kurar ve plasenta oluşumunu başlatır. Bu arada iç hücre kitlesi büyür ve farklılaşmaya başlar. Bunun bir bölümü yolk kesesi gibi başka embriyo dışı dokuları oluştururken, geri kalan kısmı diğer omurgalılardakine benzer şekilde, gastrulasyon, nörülasyon ve diğer işlemleri geçirerek gerçek embriyoyu oluşturur. Erken Memeli Embriyosu Kurallara Son Derece Uyar • İlk birkaç hücre bölünmesiyle oluşan blastomerler uyum yeteneğine sahiptir. – Başlangıç koşulları bozulsa da erken evrede gelişimsel sistem kendini düzenleyebilir ve normal yapılar geliştirir. Böyle embriyo yada embriyo parçalarına düzenleyici “regulative” denir. – İç hücre kitlesindeki her bir hücre totipotent özelliktedir ve germ hücreleri de dahil olmak üzere erişkin vücudunun herhangi bir bölümünü meydana getirebilirler. • Eğer erken embriyo ikiye ayrılırsa bir çift embriyo gelişebilir. İki hücreli embriyoda hücrelerden biri iğne ile öldürülürse diğerinden normal fare gelişebilir. • Tersine iki adet sekiz hücreli fare embriyosu tek bir morula oluşturmak üzere bir araya getirildiğinde tek normal bir birey gelişebilmektedir. Epitel ve Mezenşim Arasındaki Etkileşimler Dallanan Tübüler Yapıları Oluşturur • Omurgalılar nispeten büyük hayvanlardır ve hacimlerinin büyük bölümünü bağ dokuları oluşturmaktadır. • Diğer yandan, boşaltım, besin emilimi ve gaz alışverişi için çeşitli tipte geniş epitel yüzeylere gereksinimi vardır. Bunların çoğu, bileşik organları oluşturmak üzere epitelin embriyonik bağ dokusuna yayıldığı, dallanma morfogeneziyle oluşan tübüler yapılardır. • Akciğeri oluşturmak üzere önsindirim kanalının tabanındaki endoderm, tomurcuklanarak uzadıkça tekrar tekrar dallanan bir tüp sistemi halindeki bronş ağacını oluşturmak için, komşu mezenşimin içine doğru gelişir. • Aynı mezenşimde, endotel hücreler de (kan damarlarını kaplayan hücreler) yayılım gösterir ve böylece akciğerde gaz alışverişi için gerekli olan yakın ilişkili hava yolları ve kan damarları sistemi oluşur. • İşlemin tamamı, mezenşim ve içine doğru yayılım gösteren epitel tomurcukları arasındaki karşılıklı sinyal alışverişine bağlıdır. Bu sinyaller farelerde fibroblast büyüme etmeni (FGF) ailesine üye proteinler ve etki ettikleri reseptör tirozin kizazlar dır. • FGF10 molekülü, gelişen epitel tüplerin uç kısımlarına yakın mezenşim hücre kümelerinde ifade edilirken, reseptörü epitel hücrelerin kendisinde ifade edilir. Epitelin mezenşime yayılımı bir davetle ( FGF10’a yanıt olarak) gerçekleşmektedir. • Büyüyen epitel tüplerinin yayıldıkça dallanmasını sağlayan nedir? Zıt yönde (tomurcukların ucundaki hücrelerden mezenşime) gönderilen Sonic hedgehog sinyaline bağlı olduğu düşünülür. Sonic hedgehog sinyali tomurcuğun uzayan ucuna en yakın mezenşim hücrelerindeki FGF10 anlatımını durdurarak FGF salgılayan kümeyi iki ayrı kümeye böler ve böylece, tomurcuğun iki dala ayrılmasına neden olur. NÖRAL GELİŞİM • Sinir hücreleri (nöronlar) özelleşmiş hücreler arasında en eski olanıdır. Diğer hücrelere sinapslarla bağlanan uzun aksonları ve dentritleri bulunan bir nöron, her şeyden önce uzamış biçimiyle sıra dışıdır. • Nöral gelişimi anlama yolundaki temel mücadele, akson ve dentrit uzamasının, doğru eşlerini bulmasını ve işlevsel bir ağ oluşturmak için birbirleriyle seçici olarak sinaps yapmalarının nasıl gerçekleştiğini açıklamak içindir. • Çeşitli nöron sınıfları, glia hücreler, duyu hücreleri ve kaslardan oluşan tipik bir sinir sisteminin bileşenleri, embriyonun birbirinden uzak bir çok noktasından kaynaklanır ve başlangıçta bağlantıları yoktur. Sonraki evrede, gen düzenleyici mekanizmaların denetiminde akson ve dentritler özgül yolları izleyerek uzar ve sistemin ayrı bölümleri arasında, geçici fakat düzenli bir bağlantı ağı kurar. Erişkin yaşamına kadar süren son evrede, birbirinden uzak noktalara dağılmış bileşenler arasındaki elektrik sinyallerinin akışına bağlı olarak, bağlantı ayarları ve ince oluşumlar gerçekleşir. Doğdukları Yer ve Zamana Bağlı Olarak Nöronlara Farklı Özellikler Yüklenir • Nöronlar hemen hemen her zaman glial hücrelerle birlikte oluşur. Glial hücreler nöronlara bir destek çevre oluşturur. Her iki hücre tipi de ektodermden gelişir. Omurgalılarda merkezi sinir sistemine (omurilik, beyin ve gözün retinası) ait nöronlar ve glial hücreler, nöral tüpü oluşturmak için kıvrılan ektodermin bir bölümünden köken alırken, periferik sinir sistemine ait olanlar temelde nöral tepeciklerden türer. • Nöral tüp başlangıçta tek tabaka epitelden oluşur. Zamanla kalınlaşır ve karmaşık bir yapıya dönüşür. • Epitelin yapışkan özelliğini gila hücreleri sürdürür ve kalınlığı boyunca uzanan bir yapı iskelesi oluşturur. • Yeni doğan nöronlar bu glia hücreleri arasında göç eder, varış yerinde olgunlaşır ve akson ve dendritlerini uzatır. • Nöral tüpün ventral ve dorsal tarafından salgılanan sinyal ileti proteinleri, birbirine zıt morfogenler gibi davranarak, farklı dorsoventral düzeylerde oluşan nöronların farklı gen düzenleyici proteinleri antamasına yol açar. Baş kuyruk ekseni boyunca da Hox genlerinin anterioposterior anlatım örüntüsünü ve diğer morfogenlerin etkinliğini yansıtan farklılıklar bulunmaktadır. • Nöronların oluşumu merkezi sinir sisteminin her bölgesinde günler, haftalar, hatta aylar sürer ve bu, hücreler farklı karakterleri doğum tarihlerine göre edindikleri için, daha da büyük çeşitlilik yaratır. Doğumda Bir Nörona Yüklenen Nitelik Oluşturacağı Bağlantıyı Belirler • Gen anlatımındaki farklılıklar nöronların niteliğini ayarlar ve farklı eşlerle bağlantılar kurmalarını sağlarlar. Örn. Omurilikte ventralde yerleşim gösteren hücre kümeleri, Islet/Lim homeokutu ailesinden olan genleri anlatır ve aksonlarını özgül kas alt kümeleri ile bağlantı kuracak şekilde uzatarak, motor nöronlar şeklinde gelişir. Eğer gen anlatım örüntüsü yapay olarak değiştirilirse, nöronlar farklı hedef kaslara uzantı yapar. • Farklı varış noktaları, aksonların sinir hücresinin gövdesinden dışarı doğru büyüyerek yaptığı farklı yolak seçimlerini ve yolculuklarının sonunda farklı hedef hücrelerini seçici olarak tanımalarını yansıtır. – Omuriliğin dorsal bölümünde, vücut çevresinden gelen duyusal bilgileri alan aktaran nöronlar bulunmaktadır. Bunların bazıları aksonlarını dorsal tarafa gönderirken, diğerleri ventral tarafa gönderir; bazıları baş tarafına gönderirken, kuyruk ve yine bazıları nöral tüp tabanından geçip vücudun diğer tarafına gönderir. Her Akson veya Dentrit Uçtaki Büyüme Konisi Aracılığıyla Uzar • Nöronun akson ve dentrit uzantıları büyüme konisiyle gerçekleşir. • Büyüme konisi kendini çevreleyen dokular arasından geçerken, ince yapılı dentrit ve aksonları arkasından sürükler. • Nöronlar, başlangıçta hepsi birbirine benzeyen uzantılarını gövdelerinden çıkarmaya başlar; büyüme konilerinden birinin hızını aniden arttırdığı gözlenir ve aksona özgü proteinleriyle kendi kendini akson olarak belirler. • Sinir hücresinden gelişen uzantıdaki büyüme konisi, önünde uzanan alanı filipod ve lamellipodlarla sürekli inceleyerek günde 1 mm hızla ileriye doğru uzar. Büyüme Konisi Tanımlanmış Bir Yol Boyunca in vivo Gelişmekte Olan Nörite Klavuzluk Eder • Hayvanlarda büyüme konisi yolunu bulabilmek için çok sayıda uyarıyı kullanarak hedefine doğru yolculuk eder. – Daima üzerinde sürüklenebileceği bir hücre dışı matris dokusuna veya hücre yüzeyine gereksinim duyar. – Büyüme konileri genellikle diğer nöritlerin öncülük ettiği rotaları temas yoluyla izler. – Hücre hücre yapışma molekülleri aracılık eder. [immünoglobulin üst ailesinin (N-CAM) ve Ca+2 bağımlı kaderin ailesinin üyeleri]. Her iki ailenin de üyeleri genellikle aksonların büyüme konileri ve konilerin üzerinde sürüklendiği merkezi sinir sisteminin glia hücreleri, vücut periferindeki kas hücreleri, diğer çeşitli hücre tiplerinin yüzeyinde vardır. • Ardışık farklı çevresel uyaranlar da büyüme konilerine klavuzluk eder. Stratejik noktalardaki hücrelerden salgılanan kemotaktik etmenler tarafından oluşturulurlar. Örn. Komissura aksonu vücudun bir yanından diğerine geçer. Büyüme konileri taban düzleminden geçer ve dik açıyla dönüp taban düzleminin paralelinden beyin yönüne boylamsal bir rota izler. • İlk evre taban düzleminde salgılanan netrin proteininin derişimine bağlıdır. İkinci evrede taban düzleminden geçerken, yine taban hücreleri tarafından salgılanan itici Slit proteininin reseptörü Raundabout’un anlatımı artar. Raundabout’a bağlanan Slit, hücrelerin taban düzlemine girmesini engellyen bir itici olarak etkinlik göstermekle kalmaz, aynı zamanda çekici netrin molekülüne yanıt verilmesini engeller. Aynı zamanda büyüme konileri nöral tüpün yan duvarındaki hücrelerden salgılanan diğer bir itici protein olan semaforin reseptörünü de anlatmaya başlar. İki itici sınır arasında yakalanan büyüme konileri, orta hattan geçip, sıkı lif izlerinin içinden beyne doğru yol alır. commissuraless, büyüme konilerini bu sinyale karşı sağırlaştırmak için, başlangıçta Raundabout anlatımını engeller. Komissura büyüme konileri, comissuraless proteini bulunması durumunda orta hatta ilerler; orta hattın içinden geçerken comissuraless proteininin sağırlaştırıcı etkisi kaybolur ve koniler itilmeye başlar. Diğer taraftan çıktıklarında yüzeylerinde işlevsel Raundabout bulunduğundan, tekrar girişleri engellenir. Hedef Dokular Sinir Hücresi Büyümesini ve Sağkalımını Denetleyen Nörotrofik Etmenler Salgılar • Sonunda hedef bölgeye ulaşan akson büyüme konileri, burada durur ve sinaps yapar. Sinapslar genellikle aksondan dentritlere ve veya kaslara, tek yönde sinyal gönderir ancak, gelişimsel iletişim iki yönlüdür. Hedef dokudan gelen sinyaller büyüme konilerinin nereye sinaps yapacağını değil, aynı zamanda enervasyon yapan nöronlardan kaçının hayatta kalacağını da belirler. • Omurgalıların merkezi periferik sinir sistemindeki pek çok türden nöron fazladan üretilir. Hedef hücreye ulaştıktan sonra %50 veya daha fazlası ölür. • Nöronların bu yüksek orandaki ölümü bir rekabetin sonucudur. Her hedef hücre , o hedefe enervasyon yapan nöronların sağkalımını sağlayan özgül nötrofik etmenleri sınırlı miktarda salgılar. Eğer, hedef hücre miktarı arttırılırsa (uzuv ekleme) uzva enervasyon yapan nöronların büyük bir kısmı hayatta kalır. Tersine, uzuv çıkarılırsa nöronların hepsi ölür.. Nöronal Özgüllük, Düzenli Nöral Haritaların Oluşmasına Kılavuzluk Eder • Aynı türden olup farklı yerde bulunan nöronlardan köken alan aksonlar, yolculuk için bir araya gelir ve hedefe sıkı demetler halinde ulaşırlar, orada tekrar dağılıp, hedef sınırları içindeki farklı noktalarda sonlanırlar. • Retinadaki görsel bilgiyi beyne taşıyan nöronlar retina gaglion hücreleri olarak adlandırılırlar. Her biri görme alanının farklı bir bölgesindeki bilgiyi aktaran, bir milyondan fazla retina gaglion hücresi bulunmaktadır. Aksonları yakınlaşarak gözün optik sinir başında toplanır ve optik sap boyunca beyne kadar uzanır. Memeliler dışındaki çoğu omurgalıda ana sonlanma noktası ortak beyinde geniş bir hücre alanı olan optik rektumdur. Retinal aksonlar tekrar nöronlara bağlanırken, hücre gövdelerinin retinadaki düzenine göre, öngörülebilir bir örüntüyle dağılırlar. Retinadaki komşu gaglion hücreleri, tektumdaki komşu hedef hücrelere bağlanır. Düzenli uzantılar tektumda bir görüş alanı haritası oluşturur. • Bu türden düzenli haritalar pek çok beyin bölgesinde bulunmaktadır. Örneğin işitme sisteminde, nöronlar kulaktan beyne tonotopik bir düzende uzanır ve oluşan haritada farklı perdeden sesler hakkında bilgi alan beyin hücreleri, piyanonun tuşlarına benzer şekilde , bir hat üzerinde dizilir. Somato duyu sisteminde, dokunma duyusuyla ilgili bilgi taşıyan nöronlar beyin korteksinde bir “homunkulus(vücut yüzeyinin küçük ve biçimi bozulmuş iki boyutlu görüntüsü) haritası oluşur. Sinaptik Bağlantıların dağınık Örüntüsü Etkinlik Bağımlı Yeniden Modelleme ile Keskinleştirilmektedir • Normal bir hayvanda, retinotektal harita başlangıçta bulanık ve düzensizdir. Harita daha sonra sinaps elenmesi yoluyla netleştirilir. • Retinanın ayrı bölgelerinden aynı tektum hücresine sinaps yapılan yerlerde, aksonlardan birinin yaptığı bağlantıyı eleyen bir rekabet gerçekleşir. Ancak, retinada birbirine komşu hücrelerden gelen aksonlar işbirliği yaptığı için ortak tektum hücresindeki sinapslarını sürdürürler. Deneyim Beyindeki Sinaptik Bağlantıların Örüntüsünü Şekillendirir • “Birlikte ateşleyen nöronlar, birlikte bağlantı yapar.” Olgusu, benzer şekilde, gelişmekte olan insan beyninin deneyimlerin ışığında organize olmasına da yardım eder. • Bağlantı örüntülerinin korunması, yaşamın ilk yıllarındaki deneyimlere bağlıdır. Eğer belli bir kritik dönem boyunca (insanda yaklaşık beş yaşında sonlanır) bir göz, görsel uyarılardan yoksun bırakılacak şekilde belli bir süre kapatılıp diğer gözün olağan uyarıları almasına izin verilirse, uyarı almayan göz kabukla olan bağlantısını yitirir ve tersinir olmayan bir şekilde, neredeyse tamamen kör olur. Ateşleme kuralı ile uyumlu olarak, bu durumda bir rekabet olduğunu ve görsel kabuğa sinaps yapan etkin olmayan aksonlar elenirken, etkin aksonların yaptığı sinapsların pekiştiğini varsayabiliriz. Bu yolla kabuk bölgesi bilgi taşıyan aksonlara ayrılır ve durağan olanlar için ziyan edilmemiş olur. BİTKİ GELİŞİMİ • Bitkiler ve hayvanlar yaklaşık 1,5 milyar yıllık evrimsel geçmişle birbirlerinden ayrılırlar. Çok hücreli organizasyonlarını bağımsız olarak, fakat aynı başlangıç donanımını kullanarak evrimleştirdiler. • 1) Enerjilerini diğer canlıları yiyerek değil, güneş ışığından alırlar. • 2) Hayvan hücreleri gibi hareket etmelerini engelleyecek şekilde yarı sert bir hücre duvarı ile kaplanmış ve birbirlerine yapıştırılmıştır. • Çoğu bitkinin gelişimi çevreden ciddi şekilde etkilenir. Hareket ederek uygun çevreyi bulamayacakları için, gelişim yönlerini değiştirerek uyum gösterirler. Yaklaşımları fırsatçıdır. Toprağa dikilen bir begonya yaprağı bir kök sürebilir;kök bir filizi geliştirebilir; filiz güneş ışığında, yaprakları ve çiçekleri oluşturabilir. • Erişkin bir bitki, küçük bir standart modül setinin bir çok kopyasından oluşmuştur. • Her modül, bir sap, bir yaprak ve meristem içeren bir tomurcuktan ibarettir. • Tomurcuk yaprağın saptan ayrıldığı dallanma noktası olan, düğümde oluşur. Arabidopsis, Bitki Moleküler Genetiğinde Model Organizma Olarak Kullanılır • Çeşitliliklerine rağmen çiçekli bitkilerin ortaya çıkışı nispeten yenidir. (125 milyon yaşında; omurgalılar 350 milyon yaş). – Bir hayvanda iki kafa gelişimine neden olan mutasyon genellikle ölümcüldür, bitkide çiçek yada dal sayısını iki katına çıkaran mutasyon ölümcül değildir. • Drosophila ve Caenor haptidis elegans gibi küçüktür, çabuk üretilir ve genetik çalışmalar için uygundur. – Petri kaplarında ya da küçük saksılarda üretilir – 8-10 haftada tohum alınabilir. – Hermafrodittir. Embriyonik Gelişim Bir Kök Sürgün Ekseninin Oluşturulmasıyla Başlar ve Tohum İçinde Son Bulur • Yüksek bir bitkinin döllenmiş yumurtası, asimetrik olarak bölünmeye başlar. – Ürünlerinden biri, gerçek embriyoyu oluşturacak olan, yoğun sitoplazmalı küçük bir hücredir. – Diğeri, büyük kofullu bir hücredir ve daha da bölünerek askı denilen ve bir bakıma memelilerdeki göbek bağıyla kıyaslanabilen bir yapı oluşturur. – Askı, embriyoyu yanındaki besleyici dokuya bağlar ve besinlerin taşınmasını sağlar. Bitkinin Bölümleri Meristem Tarafından Ardışık Olarak Oluşturulur • Erişkin bitkinin bölümleri, bitkinin yanlarına yeni yapılar eklemek üzere çoğalan hücre grupları tarafından ardışık olarak yapılır. • Bu çok önemli hücre grupları tepe meristemi olarak adlandırılır. – Her meristem kendini yenileyen bir kök hücre topluluğundan oluşturur. – Bunlar bölündükçe arkalarında, meristem bölgesinden çıkıp gelişen ve nihayet farklılaşan bir döl izi bırakır. • Tepe mersitemleri dışındaki birçok hücre de, daha ileri çoğalma ve meristem potansiyelini koruma kapasitesine sahiptir. – Uzun ömürlü bitkiler kök ve gövde çevrelerini bu şekilde arttırırlar. – Bir yaralanma olunca da sürgün verebilirler. • Kök ve sürgün tepe meristemleri embriyoda zaten rudimenter olarak belirlidir. • Filizlenme sırasında tohum kabuğu parçalanır parçalanmaz meristem dışı hücrelerde, önce toprağa hemen tutunabilmek için bir kökün, daha sonra bir sürgünün belirmesini sağlamak üzere, çarpıcı bir büyüme görülür. • Tepe meristemlerindeki hızlı ve sürekli hücre bölünmeleri de buna eşlik eder. Fidenin Gelişimi Çevresel Sinyallere Bağlıdır • Filizlenmeyi takiben bitki gelişimi çevreden gelen sinyallerden kuvvetle etkilenir. • Sürgün toprak içinden hızla yolunu açarak yukarı çıkmalı ve ışığa ulaşır ulaşmaz, çelenklerini açarak fatosenteze başlamalıdır. • Yer altındaki hızlı filizlenmeden ışıkta büyümeye geçişin süresi genetik olarak programlanamaz, çünkü tohumun gömüldüğü derinlik öngörülemez. • Fideyi etkileyen ışıkla brassinoidler denilen bitki hormon grubunun yapımını baskılatarak filizlenme olur. – Brassinoid sinyalinin üretimi ya da algılanması için gerekli genlerdeki mutasyonlar, henüz karanlıktayken fide sapının yeşermesine, uzamasının yavaşlamasına ve çeneklerinin zamansız açmasına neden olur. Her Yeni Yapının Şekillenmesi, Yönlendirilmiş Hücre Bölünmesine ve Genleşmesine Dayanır • Hücre duvarları içerisinde hapis olan bitki hücresi şişebilir, gerilir ve bükülür. Gelişen bitkinin morfogenezi, geniş yönelimli hücre genleşmelerinin izlediği, düzenli hücre bölünmelerine bağlıdır. • Genleşme bitki hücre duvarına dışa doğru baskı yapan, ozmotik temelli bir turgor basıncı ile sağlanır ve yönü, hücre duvarında bulunan ve genleşmeyi tek eksene zorlayan selüloz liflerinin yönelimleriyle belirlenir. • Selülozun yönelimi de plazma zarının hemen altında bulunan ve selüloz birikimini yönlendirdiği düşünülen mikrotübül dizilerinin yönelimiyle denetlenir. • Bu yönelimler, etilen ve ciberelik asit gibi bitki büyüme düzenleyicileri tarafından hızla değiştirilebilir. Her Bitki Modülü Meristemdeki Mikroskobik İlkeller Takımından Gelişir • Sürgün tepesinde bir mili merte veya daha küçük bir alanda merkezi kısa bir kubbe görülür. • Ortadaki kubbe tepe meristemi, çevreleyen şişliklerin her biri ilkel bir yapraktır. • Bu küçük bölgede, bir kaç tam modülün henüz belirmiş , gelişmemiş bir formu bulunmaktadır. • Bitki modüllerini seri şeklinde düzenlenmesi sürgün tepesindeki olaylarla denetilmektedir. • Bu temel tema üzerindeki değişiklikler, saçaklar, dikenler, dallar ve çiçekler gibi, çok daha karmaşık yapılanmalara olanak verir. • Bitki sürgün tepesinde farklı gen setlerini açık duruma getirerek farklı uzaysal örüntüde farklı ilkel tipleri oluşturabilir. Hücre Sinyal İletimi Meristemin Sürekliliğini Sağlar • Tepe meristemi kendi devamlılığını nasıl sağlar? • Meristem hücreleri, bir bitki büyürken haftalarca, yıllarca ve hatta asırlarca kendilerini yenileyerek ve farklılaşarak yavrular üreterek çoğalmayı sürdürmelidir. • Bütün bu olaylar sırasında meristemi oluşturan hücre kümesinin boyutu hemen hemen sabit kalır. – (Arabidopsis’de 100 hücre) • WUSCHEL genindeki mutasyon tepe meristeminin yok olmasına, CLAVATA grubu genlerdeki mutasyonlar meristemin anormal büyüklükte olmasına neden olur. Bu genler meristem dokusundaki farklı hücre katmanlarında anlatım yapar. Uzun Erimli Hormon Sinyalleri Bitkinin Ayrı Bölgelerindeki Gelişim olaylarını Eşgüdümler • Bir eksen tomurcuğunun kaderi – 1) genler – 2) çevresel koşullarla belirlenir. • Bir dal ucu kopartılarak yanlara doğru büyüme uyarılabilir. • Tepe meristeminin uzaklaştırılması, uyuyan eksen meristemleri üzerindeki engellemeyi ve yeni ince dalların oluşumuna olanak verir. • Altı bitki büyüme düzenleyicisi (hormonu) oksin, ciberelinler, sitokininler, absisik asit, etilen gazı ve brassinoidlerdir.