3.BÖLÜM VİRÜS-KONAK HÜCRE İLİŞKİ TİPLERİ
advertisement

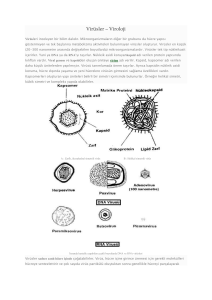
3.BÖLÜM VİRÜS-KONAK HÜCRE İLİŞKİ TİPLERİ • Üretken ilişki; Bir virüsün hücreye tutunması,girmesi ,replike olması,kendine benzeyen virüs partikülleri(progeni) sentez etmesi ve progeni virüslerin hücreden çıkarak yeni virüsleri enfekte etmesidir. • Üretken olmayan ilişki; Eğer virüs konak hücreye tutunamıyorsa veya hücreye girdikten sonra viral genler eksprese edilemiyorsa progeni sentezi gerçekleşmez. • Abortif enfeksiyon; bazı virüslerin hücre içine girmesine,nükleik asitlerinin replikasyon bölgesine taşınmasına ve hatta bazı gen ürünlerinin sentez edilmesine rağmen tüm virüs partikülü oluşmamaktadır. • Kronik enfeksiyon; bazı virüslerin replikasyonu ise hücrede kısıtlı-sınırlı olarak gerçekleşir.(semi-permissive) hücrede uzun süre kalırlar. Hücrede oluşan progeni virüs miktarı az, yavaş ve aralıklıdır; ancak viral genom hücrede bulunmaktadır. • Transformasyon; Viral DNA, hücre kromozomuna entegre olur ya da çekirdekte epizomal formda kalır ve sadece bazı erken viral ürünlerin sentezi gerçekleşir. Erken gen ürünleri, normal hücrenin fenotipik ve genotipik değişikliğe uğramasına(transformasyon) yani kanser hücresine dönüşmesine yol açar.(onkojenik transformasyon) • Latent enfeksiyon; bazı virüsler (herpes) vücuda girdikten sonra belirli hücrelerde sessiz kalırlar ve DNA ları hücre çekirdeğinde epizomal olarak replike olmadan durur, belirli faktörler varlığında aktif olur ve litik enfeksiyon oluşturur ve vücuttan temizlenemezler. VİRÜSLERİN REPLİKASYON DÖNGÜSÜ Üretken ilişki sonucunda tek bir virüs partikülünden 10-10 progeni virüs sentez edilmektedir. Bir virüsün konak hücreye girmesinden, progeni virüslerin oluşmasına dek gerçekleşen olaylar virüsün replikasyon döngüsü olarak adlandırılır; 1. Hücreye tutunma(attachment) 2. Hücre içine giriş(penetrasyon) 3. Kapsidin soyulması(uncoating) 4. Biyosentez(transkripsiyon,translasyon,replikasyon) • Erken mRNA sentezi • Erken proteinlerin sentezi • Nükleik asitlerin sentezi • Geç mRNA sentezi • Geç proteinlerin sentezi 5. Viral ürünlerin bir araya toplanması(assembly) 6. Olgunlaşma ve salınım(maturation) Hücreye tutunma(adsorbsiyon) • Eğer konak hücre virüs için özgüllük taşıyorsa hücre membranı üzerinde virüsün yüzey antijeni ile birleşebilecek en az 10-10 reseptör içerir. • Hücre yüzey reseptörleri hücrelerin sinyal iletimi, adezyon, iyon taşınması gibi fonksiyonlar için gerekli. • bu yapılar protein, karbonhidrat, glikolipid, proteoglikan yapıda olabilirler. • Tutunma elektrostatik kuvvetlerle olur. Tutunma için ligand olarak çıplak virüsler bunun için kapsid proteinlerini, zarflı olanlar ise glikoproteinlerini kullanır. Reseptör varlığı doku tropizmini belirler. • Bir hücrenin yüzeyinde farklı virüs grupları için farklı reseptörler bulunur, aynı reseptör farklı virüsler tarafından da kullanılabilir. Bir virüs birden fazla reseptöre de ihtiyaç duyabilir. Hücre içine giriş(penetrasyon) Enerji gerektirir. Çeşitli mekanizmalarla gerçekleşir. İlk ikisi hayvan virüslerinin en çok kullandığı mekanizmalardır; 1. Reseptöre bağımlı endositoz 2. Füzyon 3. Hücre membranında kapsid soyulması 4. Sinsityum oluşumu 5. Virolojik sinaps oluşumu 6. Translokasyon 1. Reseptöre bağımlı endositoz Tüm çıplak virüsler ve bazı zarflı virüsler hücretye pH a bağımlı olarak gerçekleşen endositoz yoluyla girer. Virüs gibi küçük partiküllerin endositozuna pinositoz denir. Pinositoz memelilerde klatrin, kaveola ya bağımlı ya da ikisindende bağımsız gerçekleşebilir. Bir virüsün hangi yolu kullanarak hücre içine gireceği reseptörün hangi yapıda yer aldığına bağlıdır. 2. Füzyon Zarflı virüsler hücre içine bu yol ile girerler. Viral zarf ile hücre membranı arasında kaynaşma sonucu oluşan füzyon, virüsün zarf yapısında yer alan glikoproteinler sayesinde gerçekleşir. Çıplak virüsler bu yolu kullanamaz. Bazı virüsler özel pH koşullarına ihtiyaç duyarken bazıları duymaz. 3.Hücre membranında kapsid soyulması Kapsid soyulması hücre membranına adsorbe olduktan sonra konak hücre enzimleri tarafından parçalanarak yapılır. Bu şekilde sadece nükleik asit doğrudan sitozole alınmış olur. 4.Sinsityum oluşumu Enfekte hücre ile enfekte olmayan hücrenin kaynaşması sonucu virüs bir hücreden diğerine geçebilir. 5.Virolojik sinaps oluşumu Sinaps hücreler arasında füzyon olmaksızın temas sonucu gerçekleşir ve viral ürünler enfekte hücreden sağlıklı hücreye geçer.LFA-1, ICAM-1, MTOC gibi moleküller viral yapıların geçişini kolaylaştırır. 6.Translokasyon Bir bütün halinde doğrudan penetrasyon ile hücre zarından sitoplazma içine alınırlar.(özellikle bitki virüsleri) Kapsidin soyulması(uncoating) • Kapsidin kısmen ya da tamamen ortadan kalkması ve genomun serbest kalması olayıdır. Viral nükleik asidin replike olacağı bölgeye gitmesi için öncelikle kapsid proteinlerinden kurtulması gerek. • Hücreye endozomla giren virüslerin soyulması endozom içindeki lizozomal enzimler tarafında, hücreye doğrudan giren nükleokapsidlerin ise hücre içindeki proteolik enzimler tarafından sağlanır. Çift kapsid içeren virüsler(reovirüs) ise tamaemn kapsidden soyulmadan mRNA sentezi yapılır. • Çekirdekte replike olanDNA virüsleri ise çekirdeğe hücre iskeleti aracılığı ile taşınır. Biyosentez Viral biyosentezin gerçekleşeceği hücre bölgesi virüsün nükleik asit içeriği tarafından belirlenir. Genel olarak bütün DNA virüsleri (poxyvirüsler hariç) DNA larını hücre çekirdeğinde, proteinlerini ise stoplazmad asentez ederler. Bütün RNA virüsleri de nükleik asitlerini ve viral ürünlerini hücre stoplazmasında sentez ederler. DNA virüslerinin hücre çekirdeğinde replike olmalarının nedeni , DNA kalıbından erken mRNA transkripsiyonu için hücresel DNA ya bağımlı RNA polimeraz enzimine gereksinim duymalarıdır. • Erken mRNA sentezi ; ürünleri yapısal olmayan proteinlerdir. Bu proteinler genellikle enzim aktivitesi gösteren ve virüsün nükleik asit replikasyonunda rol oynayan ürünlerdir.Nükleik asit sentezi başlamadan önce sentez edilen tüm mRNA lar ve bunların ürünleri erken olaraak adlandırılır. • Erken proteinlerin sentezi; işlenmiş mRNA lardan sitozolde yapılır. • Nükleik asitlerin sentezi; • Geç mRNA sentezi; viral nükleik asit sentezi başladıktan sonrasentez edilen tüm mRNA lar geç diye adlandırılır. • Geç proteinlerin sentezi; geç proteinler virüsün yapısal (öz yapı, kapsomer, peplomer) proteinlerdir. Biyosentez tamamlandıktan sonra oluşan viral ürünler replikasyon bölgesinde birikmeye başlar. Hücre çekirdeği ya da sitoplazmasında biriken viral nükleik asitve proteinlerin oluşturduğu bölgeler yoğun yapıların ortaya çıkmasına neden olur. Bunlara inklüzyon cisimciği denir. Nükleik asitlerin bulunduğu bölgeler bazik,proteinlerin bulunduğu bölgeler asidik boyalarla boyanabilirler. Viral nükleik asitlerin replikasyon stratejileri viral genomun yapısı, gen transkripsiyon şekli ve virion içi enzim varlığı dikkate alınır. Sınıf 1; çift iplikli DNA virüslri Çift iplikli DNA virüsleri semikonservatif olarak replike olurlar. Replikasyon öncesi DNA molekülünün her iki iplikçiği de mRNA için kalıp görevi görür. A)Adenovirüs replikasyonu B)Herpesvirüs replikasyonu C)Polyoma ve papillomavirüs replikasyonu D)Poksvirüs replikasyonu Sınıf 2 ; tek iplikli DNA virüsleri defektif ve defektif olmayan parvovirüsler bulunur. Defektif olamayanlar otonom şekilde replike olurlar , konak hücre enzimlerini kullanırlar. Dedektif olanlar replikasyon için bir başka virüsün yardımına gerek duyarlar. Sınıf 3; çift iplikli RNA virüsleri Çift iplikli 10-12 parçalı RNA içeren reovirüslerin replikasyon şeklidir. Her çift iplikli RNA segmentinin negatif iplikçiği kalıp olarak kullanılır ve tek iplikli pozitif RNA sentez edilir. Bunlar hem mRNA görevi yaparalr, hem de progeni RNA için kalıp görevi görürler. Sınıf 4 ; tek iplikli pozitif polariteli RNA virüsleri Parental RNA larını mRNA olarak kullanırve pozitif iplikçikten poliribozozmlarda doğrudan protein sentezi yapılır . Bu oldukça büyük bir proteindir. Fonksiyonel olması için proteazlarla kesilmesi gerekir. Sınıf 5; tek iplikli negatif polariteli RNA virüsleri Aynı negatif kalıp üzerinde birden fazla pozitif iplik sentez edilebilir. Yeni sentez edilen pozitif iplikler hem mRNA görevi görürler, hem de yeni negatif ipliklerin sentez için kalıp işlevi yaparlar. Bu pozitif iplik, aynı anda birden fazla progeni RNA sentezini yönetebilir. Sınıf 6; ters transkriptaz içeren tek iplikli pozitif polariteli RNA virüsleri Virion içinde RT den başka integraz ve proteaz enzimleri de bulunur. RT enzimi çok fonksiyonlu bir enzim olup, ters transkriptaz, endoribonükleaz(Rnaz-H) ve DNA polimeraz aktivitelerine sahiptir. Sınıf 7 ;terz transkriptaz içeren çembersel çift iplikli DNA virüsleri Bu grupta RT içeren, çembersel çift iplikli DNA ya sahip hepatit B virüsü yer alır. Çift iplikli çembersel DNA sının küçük bir bölümü telk iplikten oluşan bir aralık bölgesi (gap) içerir. Viral ürünlerin bir araya toplanması(assembly) Viral ürünlerin bir araya gelmesi çıplak virüslerde nükleo kapsid zarflı virüslerde ise buna ek olarak zarf yapısının oluşumunu kapsar. DNA virüslerinde tüm virüs partikülünün oluşması, çekirdekte, RNA virüslerinde ise stoplazmada olur. Kapsidin yapılanması, kapsomer ünitelerinin hidrofobik ve elektrostatik bağlarala bağlanmasıyla oluşur. ikozahedral simetrili virüslerin birçoğu önce boş kapsidi oluşturmakta, genomu sonradan kapsid içine yerleştirilmektedir. Heliksel simetrili RNA virüslerinde kapsid oluşumu ile genom paketlenmesi birlikte yapılır. Çift iplikli DNA virüsleri genomlarını paketlemek için çoğunlukla histon-benzeri molekülleri kullanır. Bunlar virüse ait olabileceği gibi virüsün hücreden aldığı hücresel histon molekülleri olabilir. Olgunlaşma ve salınım(maturation Paketlenme ve progeni virionların oluşmasından sonra salınım aşaması 3 yolla gerçekleşir; a)Hücre lizisi Çıplak virüsler hücrenin erimesine yol açarak(otolizis) hücreden dışarı çıkarlar. b)Tomurcuklanma ve eksositoz Zarflı virüsler hücreden tomurcuklanma ile çıkar ve membran yapısından bir kılıf almadan önce sentezledikleri peplomerleri membrana birleştirerek kendilerine özgül bir antijenik yapı kazandırırlar. Sitoplazmik membrandan tomurcuklananlar doğrudan dış ortama salınır, çekirdek, ER ve golgi gibi hücre içi organellerden tomurcuklanan virüsler ise ekzositoz ile hücre dışına çıkarlar. Tomurcuklanma sırasında hücre çok az zedelenebileceği gibi , bazılarının tomurcuklanması hücrede ciddi zedelenmeye neden olur. c)Aktin kuyrukları ile dışarı itilme Poxyvirüslerin hücreden çıkışı bu yolla olur. Sitoplazmada çoğalarak sitozolde biriken poxyvirüsler golgi sistemine taşınır ve zarflarını buradan alırlar. Hücre içi virüs partiküllerinin bir kısmı hücreden ayrılamaz aktin filemant yardımıyla itilerek hücreden ayrılır. Hücresel zarların yapısında bulunan ve raft olarak adlandırılan kolesterol ve sfingolipidden zengin hareketli özel yapılar,sinyal iletimi ve endositoz gibi fonksiyonlarının yanı sıra viral yaşam döngüsünde de rol alırlar. Viroporinler hücre membranında porlar oluşturur virüs salınımı kolaylaşır.











![(Microsoft PowerPoint - Viral_replikasy\375on [Uyumluluk Modu])](http://s1.studylibtr.com/store/data/000866796_1-a2dd980230ca32afc0bef516628ca139-300x300.png)








